Análisis de la expresión de mRNA de Cx43 en células de la granulosa de pacientes sometidas a tratamiento de reproducción asistida
P. Sánchez-Aparicio1, C. Romero2, J.M. Cuadros-Fernández3, J. Corredera2, T. Muñoz2, E. Fernández2, B. Hurtado de Mendoza2.
1Departamento de Biología Celular y del Desarrollo, CIB, CSIC. Madrid. 2Instituto Ginecológico La Cigüeña. Madrid. 3Clínica de Medicina de la Reproducción y Ginecología FivMadrid. Madrid.
Email:aparicio@cib.csic.es
La foliculogénesis es un proceso complejo y estrechamente regulado. La coordinación entre el oocito y las células de la granulosa en los folículos ováricos asegura el proceso de maduración folicular y está mediada por las uniones tipo gap o gap junctions. Trabajos llevados a cabo en animales de experimentación demuestran que la Conexina 43 (Cx43), proteína que forma parte de las GJ, juega un papel esencial en el proceso de la foliculogénesis. Aunque se tiene un amplio conocimiento de este proceso, existen algunos aspectos del desarrollo folicular aún desconocidos; por ejemplo , si posibles alteraciones en la regulación transcripcional de Cx43 podrían llegar a comprometer el proceso de maduración folicular en humanos. El objetivo de este trabajo es determinar si las células de la granulosa de pacientes con trastornos de infertilidad son capaces o no de expresar Cx43. La expresión de Cx43 a nivel de mRNA se analizó por técnicas de RT-PCR en las células de la granulosa de las pacientes, tras punción folicular. Nuestros resultados muestran que en todos los casos pudimos detectar la presencia de mRNA de Cx43, independientemente del diagnóstico previo o de la calidad oocitaria. Sin embargo, dada la extrema sensibilidad de la técnica para detectar incluso ínfimas cantidades de mRNA de Cx43, no podemos descartar que existan diferencias significativas no detectadas en los niveles de expresión de Cx43 entre los distintos casos analizados. Asimismo, se discuten otras posibles razones que justifiquen los resultados obtenidos.
INTRODUCCIÓN
La producción de gametos funcionales es esencial e imprescindible para la transmisión del genoma a las generaciones sucesivas. Los gametos haploides dan lugar a un zigoto tras la fecundación y luego a un embrión cuyo proceso de desarrollo es el resultado de la puesta en funcionamiento del programa genético contenido en su genoma. El proceso de formación de los gametos, depende de su interacción con las células somáticas circundantes, lo que resulta crucial para la fertilidad.
El ovario es una estructura dinámica en la que los folículos ováricos están en contínuo desarrollo a partir del estadio primordial, asegurando la producción exitosa de gametos femeninos. De hecho, el ciclo ovárico constituye un ejemplo excelente de fenómenos repetidos de proliferación celular, diferenciación y apoptosis, que ocurren de una manera cíclica, regulada y perfectamente sincronizada (Guthrie and Garret, 2001; Vanderhyden, 2002; Van den Hurk and Zhao, 2005; Hussein 2005).
Los folículos primordiales se forman durante el desarrollo prenatal y están constituidos por un oocito detenido en la profase de la primera división meiótica y una capa de células somáticas denominadas células de la granulosa. El proceso de crecimiento folicular culmina en el momento de la ovulación, coincidiendo con la iniciación de los ciclos reproductivos.
Durante la foliculogénesis, los oocitos avanzan en su proceso de maduración, mientras que las células de la granulosa desarrollan capacidades esteroidogénicas específicas. Este proceso requiere mecanismos complejos de regulación que implican rutas de señalización intrínsecas y extrínsecas.
En el caso de las intrínsecas, numerosos factores de crecimiento intra-ovario, pertenecientes a distintas famillias, influyen a través de la señalización paracrina en el crecimiento y la maduración folicular (Nilssoon and Skinner, 2001; Gilchrist et al., 2004).
Por otra parte, existen numerosas evidencias que demuestran que los estadios tardíos del desarrollo folicular son dependientes de la señalización extrínseca en la que participan las hormonas hipofisarias FSH (hormona folículo-estimulante) y LH (hormona luteinizante). Especialmente la hormona FSH es el mayor promotor de la maduración folicular, fomentando la proliferación de las células de la granulosa y regulando la producción de la hormona ovárica estradiol. La hormona LH juega por su parte un papel en estadios más avanzados del desarrollo folicular, estimulando la maduración del oocito, la ovulación y posteriormente el proceso de luteinización.
La coordinación entre el oocito y las células de la granulosa está medida por la comunicación célula-célula que es generada por las uniones tipo gap o gap junctions (GJ) (Kidder and Mhawi, 2002; Matzuk et al., 2002).
Las GJ son agrupaciones de canales intercelulares que permiten una comunicación citoplasmática directa entre células adyacentes. Estos canales están a su vez formados por subunidades de proteínas denominadas conexinas (Cxs); y permiten el paso de pequeñas moléculas -iones, metabolitos, segundos mensajeros- haciendo de este modo posible la sincronización del comportamiento celular (Evans and Martin 2002; Sohl and Willecke, 2004).
La conexina 43 (Cx43) es el miembro de la familia de las conexinas que se expresa mayoritariamente en el folículo ovárico y cuyo papel en la foliculogénesis es más relevante (Juneja et al, 1999; Ackert et al., 2001). Estudios llevados a cabo con ratones knock-out para Cx43 ponen de manifiesto el papel esencial de esta molécula para mantener la comunicación entre las células de la granulosa. Los resultados obtenidos demuestran que se requiere la presencia de Cx43 para sostener la proliferación de estas células y la posterior maduracióon folicular. Además, trabajos recientes demuestran la implicación de Cx43 en el desarrollo preimplantacional de embriones humanos (Bloor et al., 2004).
La Cx43 se detecta ya en los folículos primordiales y su presencia se hace más evidente con el crecimiento folicular; su expresión está sujeta a modulación durante el ciclo ovárico (Gittens et al., 2005), estando los niveles de esta molécula afectados por las hormonas FSH y LH (Granot and Dekel, 2002). La hormona FSH estimula la síntesis de Cx43 e incrementa simultáneamente el número de canales intercelulares funcionales. Por el contrario, la hormona LH, en un respuesta inmediata, interrumpe la comunicación célula-célula, aparentemente por fosforilación y modificación conformacional de la molécula de Cx43, lo que conduce al cierre de los canales (Lampe and Lau, 2004). Este efecto inmediato de LH es seguido por una respuesta tardía, que implica la eliminación de la proteína por estimulación de la reducción de los niveles de mRNA y la desaparición final de las GJ (Granot and Dekel, 2002). La actividad biológica de la hormona FSH tiene lugar a través de la ruta de señalización PKA (proteína quinasa A) dependiente de AMP cíclico, mientras que la hormona LH ejerce su acción biológica no sólo a través de la ruta de transducción de señales PKA sino también de la PKC (proteína quinasa C). Además, como las hormonas FSH y LH estimulan la producción por parte de las células de la granulosa de estrógenos y progesterona, respectivamente, se ha planteado recientemente si serían estas hormonas las que podrían actuar como mediadores de la modulación de Cx43 inducida por FSH y LH en el ovario.
Aunque en líneas generales se tiene un amplio conocimiento sobre los procesos que conducen al desarrollo folicular, aún se desconocen algunos aspectos específicos, como las implicaciones biológicas de la comunicación celular vía GJ de Cx43 en este proceso.
Parece razonable pensar que la expresión de Cx43 en las células de la granulosa estaría sujeta a un complejo sistema de regulación transcripcional. Teniendo en cuenta el papel tan relevante que desempeña en el proceso de la foliculogénesis, cabría cuestionarse si alteraciones en la regulación transcripcional de Cx43 podrían llegar a comprometer el proceso de maduración folicular en humanos.
Con la intención de evaluar esta posibilidad, desarrollamos una estrategia de análisis dirigida al estudio de la capacidad de expresión de esta molécula a nivel de mRNA, en una muestra de pacientes con trastornos de infertilidad de más de dos años de evolución y sometidas a tratamiento de reproducción asistida por ICSI. Los primeros resultados obtenidos por técnicas de RT-PCR se presentan a continuación en este trabajo.
MATERIALES Y MÉTODOS
Células de la granulosa: Tras la punción folicular, se procedió a la limpieza de la totalidad de los oocitos extraídos en cada caso, sin excluir ninguno de ellos, aún siendo los oocitos en la observación preliminar de carácter inmaduro o incluso atrésicos. La totalidad de las células de la granulosa liberadas mecánicamente, tras un breve tratamiento con hialuronidasa, se recogió en tubos de fondo cónico de 15ml y se lavó con 10ml de PBS (tampón fosfato salino). Tras centrifugación a 1500rpm durante 20 minutos, el sobrenadante se eliminó por aspiración y el pellet se procesó para la extracción de RNA.
Células Huvec: Las células Huvec se obtuvieron por cultivo primario de células endoteliales procedentes de cordón umbilical de niños recién nacidos, y se emplearon como fuente de RNA en la puesta a punto de las condiciones de PCR de este trabajo.

RT-PCR: La extracción de RNA de las células de la granulosa se llevó a cabo con fenol/cloroformo. La totalidad del RNA precipitado se resuspendió en 9ml de H2O y se procedió a realizar una RT (Retro Transcripción) para la obtención de ADN complementario (ADNc) a partir de RH (hexámeros aleatorios) y enzima M-MLV (Promega). Las condiciones de RT se especifican a continuación: 25ºC 10´/42ºC 30´/99ºC 5´/4ºC 5´. A partir de 3ml de la muestra de ADNc se realizó una PCR (Reacción en cadena de la Polimerasa), utilizando oligonucleótidos específicos -sentido y antisentido- para Cx43 y GAPDH respectivamente y enzima Taq polimerasa (Applied Biosystem). Nosotros diseñamos los oligonucleótidos específicos para Cx43 (ver Fig. 1) y empleamos oligonucleótidos usados habitualmente por otros autores (5´-GGCTGA-GAACGGGAAGCTTGTCA-3´s y 5´CGGCCATCACGCCACAGTTTC-3’a) para GAPDH. Las condiciones de PCR se indican a continuación: 94ºC 3´/35 ciclos (94ºC 30″ – 55ºC 30″ – 72ºC 30″)/ 72ºC 5´/4ºC 5´. Las muestras amplificadas se corrigieron en gel de agarosa al 1,5% utilizando como marcador AHind III. Los geles fueron fotografiados sobre un transiluminador de luz ultravioleta.
RESULTADOS
La Tabla I recoge la información relevante sobre las primeras cuatro pacientes analizadas en este trabajo: los casos se eligieron al azar sin ningún condicionante previo entre el conjunto de pacientes con trastornos de infertilidad sometidas a tratamientos de reproducción asistida en el Instituto Ginecológico La Cigüeña. En todos los casos se trataba de pacientes sometidas a ciclos de ICSI, con dos rangos de edad bien diferenciados (<35 años/ >35años). Todas las pacientes presentaban factor femenino, mientras que, además en tres de los casos existía factor masculino adicional. Además, la tabla incluye información adicional sobre la calidad oocitaria, valorada globalmente en el conjunto de los oocitos obtenidos tras punción folicular en cada caso, así como datos sobre las gestaciones resultantes.
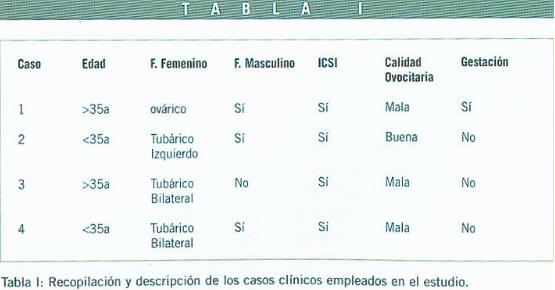
Las células de la granulosa de las pacientes analizadas se obtuvieron en número reducido tras la limpieza minuciosa de los oocitos previa a la ICSI. El procedimiento así diseñado nos aseguraba recoger exclusivamente las células de la granulosa circundantes y en contacto más íntimo con el oocito (corona radiata) y descartar de este modo otro tipos celulares que pudieran contaminar nuestro objeto de estudio. Es importante reseñar que en ningún caso se seleccionaron oocitos en función de su calidad o estado de desarrollo; es decir, se optó por limpiar de células de la granulosa todos aquellos oocitos obtenidos tras la punción folicular. De este modo obtuvimos un conjunto heterogéneo de células de la granulosa procedentes tanto de vesículas germinales como de oocitos en MI o MII, e incluso oocitos atrésicos. A partir de estas muestras se procedió al análisis de la expresión de Cx43 por RT-PCR en cada caso, con la intención de estudiar la capacidad de expresión de mRNA de Cx43 en estas pacientes y poder determinar así si los trastornos de infertilidad diagnosticados podrían asociarse a una alteración en la regulación transcripcional de Cx43.
Para poder llevar a cabo la amplificación del ADNc específico de Cx43 obtenido tras retro-transcripción (RT), diseñamos una pareja de oligonucleótidos sentido y antisentido. La Figura 1 recoge las secuencias diseñadas u tilizando las bases de datos disponibles, así como el programa OLIGO, desarrollado específicamente para el diseño de oligonucleótidos.Ambas secuencias se enmarcaban en exones diferentes para evitar así posibles contaminaciones por ADN genómico en nuestros experimentos. Además, llevamos a cabo algunos ensayos previos con RNA de células Huvec, lo que nos permitió establecer las condiciones de PCR y confirmar la especificidad de nuestros oligonucleótidos. Se utilizaron estas células como control por expresar Cx43 de forma constitutiva, obteniéndose así un producto de amplificación mayoritario correspondiente al tamaño esperado de 283 pares de bases (bp), de acuerdo al cálculo teórico según los oligonucleótidos escogidos (Fig 2).Además, para estas mismas células se obtuvo un producto de amplificación minoritario, probablemente correspondiente a una variante por splicing alternativo de Cx43. Como control positivo de la retro-transcripción empleamos una pareja de oligonucleótidos para GAPDH; el producto de amplificación en este caso fue una banda de 420bp, que por su tamaño corre retrasada respecto a la banda correspondiente al producto de 283bp específico de Cx43.
tilizando las bases de datos disponibles, así como el programa OLIGO, desarrollado específicamente para el diseño de oligonucleótidos.Ambas secuencias se enmarcaban en exones diferentes para evitar así posibles contaminaciones por ADN genómico en nuestros experimentos. Además, llevamos a cabo algunos ensayos previos con RNA de células Huvec, lo que nos permitió establecer las condiciones de PCR y confirmar la especificidad de nuestros oligonucleótidos. Se utilizaron estas células como control por expresar Cx43 de forma constitutiva, obteniéndose así un producto de amplificación mayoritario correspondiente al tamaño esperado de 283 pares de bases (bp), de acuerdo al cálculo teórico según los oligonucleótidos escogidos (Fig 2).Además, para estas mismas células se obtuvo un producto de amplificación minoritario, probablemente correspondiente a una variante por splicing alternativo de Cx43. Como control positivo de la retro-transcripción empleamos una pareja de oligonucleótidos para GAPDH; el producto de amplificación en este caso fue una banda de 420bp, que por su tamaño corre retrasada respecto a la banda correspondiente al producto de 283bp específico de Cx43.
Una vez establecidas las condiciones técnicas adecuadas, procedimos al análisis del Caso 1 (ver Tabla I). Se trata de una mujer mayor de 35 años con factor ovárico y factor masculino asociado, que se sometió a un ciclo de ICSI. La totalidad de los oocitos se limpiaron, incluyendo los atrésicos, y se obtuvo así una muestra heterogénea de células de la granulosa. Se procedió a la extracción de RNA y posteriormente a la RT-PCR para detectar mRNA específico de Cx43. Como se aprecia en la Figura 2, la detección de una banda única del tamaño esperado equivalente a 283bp demuestra la existencia de mRNA de Cx43 en las células de la granulosa de esta paciente. La ausencia de bandas adicionales asegura la especificidad de los fragmentos obtenidos.
El Caso 2 corresponde a una mujer menor de 35 años; en este caso con factor tubárico y con factor masculino adicional, que se somete a un ciclo de ICSI. Según se observa en la Figura 3, las células de la granulosa obtenidas según el procedimiento descrito previamente expresan igualmente mRNA de Cx43.

El Caso 3 es el de una mujer mayor de 35 años, con factor tubárico bilateral y sin factor masculino, que por su historia clínica se somete a un ciclo de ICSI. Los resultados obtenidos por RT-PCR se muestran en la Figura 3. Nuevamente, al igual que en los casos anteriores, las células de la granulosa expresan inequívocamente Cx43.
Por último,los resultados obtenidos para el Caso 4 (mujer menor de 35 años), que al igual que el caso anterior es un factor tubárico bilateral, aunque asociado también a factor masculino, se recogen en la Figura 3. Como se aprecia, los resultados son idénticos a los obtenidos para los tres casos anteriores, confirmándose la presencia de mRNA de Cx43 en este paciente.
La homogeneidad de los resultados obtenidos, aún a pesar de lo limitado de la muestra, sugieren que la capacidad de expresión de Cx43 a nivel de mRNA podría conservarse independientemente de la edad, factor ovárico/tubárico asociado o la calidad oocitaria global. Sin embargo, nuestros datos no nos permiten concluir que la molécula de Cx43 se esté expresando a nivel de proteína en dichas células.
DISCUSIÓN
Teniendo en cuenta el papel fundamental que la Cx43 parece desempeñar en la foliculogénesis, como indican los estudios llevados a cabo con animales experimentales, nos planteamos si posibles alteraciones en la regulación transcripcional de Cx43 podrían llegar a comprometer el proceso de maduración folicular y afectar así la calidad oocitaria también en humanos. Por ello, desarrollamos una estrategia de análisis centrada en el estudio de la expresión de Cx43 en células de la granulosa procedentes de pacientes con trastornos de infertilidad. Como primera aproximación, hemos intentado realizar un estudio cualitativo de la expresión de Cx43. De este modo, utilizamos una técnica como la RT-PCR, sensible a la detección de mínimas cantidades de Cx43.
Nuestros resultados muestran que en todos los casos analizados las células de la granulosa son capaces de expresar Cx43 a nivel de mRNA, independientemente del diagnóstico previo de los pacientes o incluso de la calidad oocitaria global.
La ausencia de correlación entre la expresión de Cx43 y la calidad oocitaria podría deberse al hecho de que las células de la granulosa se analizaron en su conjunto, formando parte de una población heterogénea; es decir,sin tener en cuenta su procedencia de oocitos maduros o atrésicos. Pudiera ser que sólo las células de la granulosa rodeando a oocitos atrésicos carecieran de Cx43. Esta circunstancia habría escapado a nuestra valoración, puesto que la técnica habría sido capaz de detectar incluso pequeñas cantidades de Cx43 procedentes de algunos de los oocitos maduros presentes en la muestra. Por tanto, no podemos descartar haber detectado la Cx43 residual procedente de algún oocito de relativa buena calidad.
Asimismo, la técnica de análisis por RT-PCR sólo permite hacer una valoración cualitativa de la expresión de Cx43. Esto significa que sólo podemos valorar si las células de la granulosa de las pacientes analizadas son capaces o no de expresar Cx43.
Sin embargo, de existir diferencias significativas en los niveles de expresión de estos pacientes, nosotros no habríamos podido detectarlas. Debido a la alta sensibilidad de la técnica utilizada, las medidas realizadas alcanzan el nivel de saturación casi de inmediato. Nosotros llevamos a cabo en todos los casos una PCR estándar de 35 ciclos, lo que justifica que se alcance siempre la saturación. Sólo la puesta a punto de una PCR cuantitativa en tiempo real, que estamos realizando actualmente, nos permitirá elucidar posibles cambios en los niveles de expresión de mRNA de Cx43 para cada una de las pacientes analizadas. En el supuesto de que existiera expresión de Cx43 en todos los casos analizados y que realmente se dieran diferencias de expresión cualitativas entre las células de la granulosa dependiendo de si rodean oocitos maduros o atrésicos, sería posible correlacionar el nivel de expresión de Cx43 en una muestra heterogénea de células de la granulosa con la calidad oocitaria global, puesto que en cada caso la contribución relativa de oocitos de buena calidad sería diferente.
Además, es interesante destacar que el hecho de detectar mRNA de Cx43 en una población celular no nos asegura que este mRNA se esté traduciendo eficientemente a proteína. Es decir, podría darse la circunstacia de que existiera un fallo a nivel postranscripcional o a nivel transduccional, de forma que aún existiendo mRNA en la célula no se sintetizase proteína. Un análisis de la expresión de Cx43 a nivel de proteína en las células de la granulosa, por técnicas de inmunofluorescencia o western-blot, serviría para este propósito. Más aún, podría darse el caso de que, incluso existiendo GJ de Cx43 entre unas células de la granulosa y otras, estas GJ no fueran realmente funcionales, impidiéndose así una correcta comunicación intercelular y una sincronización apropiada del comportamiento celular.
Así pues, este estudio pone de manifiesto la necesidad de desarrollar un análisis futuro más exhaustivo y detallado de la expresión de Cx43 a nivel de mRNA y proteína en las células de la granulosa, así como una valoración de la funcionalidad de las GJ de Cx43 que nos permitiera establecer o descartar una relación inequívoca entre las GJ de Cx43 y la maduración oocitaria. Podría ser que una alteración sustancial de la funcionalidad de las GJ de Cx43 justificara la aparición elevada y recurrente de oocitos atrésicos en algunas pacientes con trastornos de infertilidad
En resumen, en este trabajo se ha estudiado la capacidad de expresión de Cx43 en las células de la granulosa de pacientes con trastornos de infertilidad que se han sometido a tratamiento de reproducción asistida. Los resultados obtenidos muestran claramente que las células de la granulosa en todos los casos analizados son capaces de expresar Cx43 a nivel de mRNA.
El papel de Cx43 en la biología de la reproducción parece ser notable no sólo por su implicación en la foliculogénesis, sino también en el desarrollo preimplantacional de embriones humanos. Por tanto, parece razonable extender nuestro análisis a un mayor número de casos, con objeto de excluir definitivamente la posibilidad de una ausencia total de expresión de Cx43 en el entorno folicular de casos particularmente extremos. Por otra parte, de los resultados presentados aquí, se desprende la necesidad de obtener resultados concluyentes a partir de experimentos de RT-PCR cuantitativa en tiempo real. Por último, en el futuro, pretendemos extender el estudio al análisis de la expresión de Cx43 a nivel de proteína y a la determinación de la funcionalidad de las GJ por técnicas de transferencia de tinción en cultivos in vitro de células de la granulosa, para evaluar si una modificación en la funcionalidad de las GJ pudiera comprometer el éxito del proceso reproductivo.
Agradecimientos
Agradecemos al Dr. J. Rey-Campos su inestimable colaboración. Este trabajo se financió con el proyecto de investigación del Ministerio de Ciencia y Tecnología PM99-0103.
Referencias
Ackert CL, Gittens JE, O´Brien MJ, Eppig JJ, Kidder GM. Intercellular communications via connexin43 gap junctions is required for ovarian folliculogenesis in the mouse. Dev Biol 2001; 233:258-20.
Bloor DJ, Wilson Y, Kibschull M, Traub O, Leese HJ, Winerhager SJ. Expression of connexins in human preimplantation embryos in vitro. Reprod Biol Endocrinol 2004; 2:1-9.
Evans WH, Martin PEM. Lighting up gap junction channels in a flash. BioEssays 2002; 24:876-80.
Gilchrist RB, Ritter LJ, Armstrong DT. Oocyte-somatic cell interactions during follicle development in mammals. Anim Reprod Sci 2004; 82-83:431-46.
Gittens JE, Barr KJ, Vanderhyden BC, Kidder GM. Interplay between paracrine signalling and gap junctional communication in ovarian follicle. J.Cell Sci 2005; 118(pt1):113-22.
Granto I, Dekel N. The ovarian gap junction protein connexin 43: regulation by gonadotropins. Trends. Endocrinol Metab 2002; 13:310-13.
guthrie HD, Garret WM. Apoptosis during folliculogenesis in pigs. Reprod Suppl 2001; 58:17-29.
Hussein MR. Apoptosis in the ovary: molecular mechanisms. Hum Reprod Update 2005; 11(2): 162-77.
Juneja SC, Barr KJ, Enders GC, Kidder GM. Defects in the germ line and gonads of mice lacking connexin43. Biol Reprod 1999; 60(5):1263-70.
Kidder GM, Mhawi AA. Gap Junctions and ovarian folliculogenesis. Reproduction 2002; 123:613-20.
Lampe PD, Lau AF. The effects of connexin phosphorylation on gap junctional communication. Int J Biochem Cell Biol 2004; 36(7):1171-86.
Matzuk MM, Burns KH, Viveiros, Eppig JJ. Intercellular communication in the mammlian ovary: oocyte carry the conversation. Science 2002; 296:2178-80.
Nilsson E, Skinner MK. Cellular interactions that control primordial follice development and folliculogenesis. J Soc Gynecol Investig 2001; 8(1SP):S17-20.
Sohl G, Willecke K. Gap Junctions and the connexin protein family. Cardiovasc Res 2004; 62(2):228-32.
Van den Hurk R, Zhao J. Formation of mammalian oocytes and their growth, differentiation and maturation within ovarian follicles. Theriogenology 2005; 63(6):1717-51.
Vanderhyden B. Molecular basis of ovarian development and function. Front Biosci 2002; 1(7): 2006-22.