GASTRULACIÓN: PROCESO CLAVE EN LA FORMACIÓN DE UN NUEVO ORGANISMO
Carmen López-Sánchez1, Virginio García-López1, José Mijares2, José Antonio Domínguez3, Francisco M. Sánchez-Margallo2, Ignacio Santiago Álvarez-Miguel1, Virginio García-Martínez1*
1Departamento de Anatomía, Biología Celular y Zoología. Universidad de Extremadura. Badajoz
2Centro de Cirugía de Mínima Invasión Jesús Usón. Cáceres.
3Instituto Extremeño de Reproducción Asistida (IERA). Badajoz.
* email: virginio@unex.es
Tras la implantación del blastocisto en el endometrio acontecen durante el desarrollo embrionario una serie de procesos morfogenéticos de significación biológica altamente relevante. Desde un embrión bilaminar, constituido por epiblasto e hipoblasto, se configura un embrión trilaminar, constituido por ectodermo, mesodermo y endodermo, debido a un proceso denominado gastrulación, un proceso que no es exclusivo de vertebrados, basado en un proceso de división, migración y diferenciación celular. En el presente trabajo realizamos una descripción precisa del proceso de gastrulación, con especial referencia a las estructuras que sucesivamente se van formando, y analizando los posibles factores implicados en la determinación de cada uno de ellos. Rev Asoc Est Biol Rep 2013; 18(1):29-41.
1. Introducción
El embrión es transferido a la cavidad uterina en las técnicas de reproducción asistida (RA) en la etapa de blastocisto, o bien en alguna de las fases previas a su formación. El embriólogo clínico está familiarizado con las etapas del desarrollo embrionario preimplantacional, y las nuevas tecnologías, como es el caso de la embriocinética (Campbell et al., 2013), que aportan constantemente nuevos conocimientos sobre las particularidades de este proceso. En cualquier caso, el proceso de implantación comienza cuando el embrión eclosiona de la zona pelúcida, desencadenándose un complejo proceso de placentación que suele ser seguido por parte de los embriólogos, aunque de forma indirecta, por la formación del saco embrionario, que se observa ecográficamente.
Sin embargo, de forma simultánea con la implantación, mediante la invasión del endometrio por parte del trofoectodermo, la masa celular interna (MCI) del blastocisto sufre una serie de vertiginosos cambios que conllevarán finalmente a la formación del embrión propiamente dicho. Estas etapas del desarrollo que conllevan la formación de las capas germinales y si cabe más relevante aún, la formación del futuro plan corporal del organismo, están gobernadas por el proceso de gastrulación, una etapa crucial del desarrollo donde además se establece la identidad propia de cada organismo y que supone la especificación de grupos celulares para la formación de los distintos órganos.
En este artículo mostramos una visión general de los acontecimientos que tienen lugar en la MCI, desde la etapa de blastocisto hasta que el futuro organismo ya se puede identificar con una estructura básica similar a la que tendrá en etapas más avanzadas del desarrollo, incluyendo el adulto.
Conceptualmente, es un proceso transcendental pues en esta fase se consigue que desde un grupo indeterminado de células y con pluripotencialidad para originar cualquier tipo celular, se establezcan los esbozos de los futuros órganos en sus posiciones correctas y que las células se comprometan a sus destinos de forma prácticamente irreversible. Aunque las etapas previas a la gastrulación son muy diferentes entre los mamíferos y el resto de los vertebrados, debido principalmente a diferencias en el proceso de segmentación, que a su vez está determinado por el tamaño de los gametos femeninos (huevos), cuando se forma el disco embrionario y consecuentemente se inicia la gastrulación, los mecanismos de desarrollo para establecer el futuro organismo siguen patrones muy similares, independientemente del grupo de vertebrados que consideremos. Esto es un hecho relevante, denominado convergencia evolutiva, que indica la relevancia del mecanismo de gastrulación. Básicamente podríamos considerar a esta orquestada secuencia de procesos de desarrollo que ocurre durante la gastrulación como una especie de cuello de botella por el que han de pasar todos los vertebrados para conseguir la formación de un nuevo organismo. Debido a esto, gran parte de los datos más relevantes del proceso de gastrulación han sido obtenidos del estudio de animales más asequibles para la experimentación y manipulación que los embriones de mamíferos, como es el caso de los embriones de aves. Por lo tanto, gran parte de la información experimental que ofreceremos se ha conseguido con embriones de gallina, aunque es perfectamente equiparable y trasladable no sólo al resto de los vertebrados sino también y más relevantemente a la especie humana.
2. El disco embrionario: epiblasto e hipoblasto
Aunque cuando el blastocisto es transferido la MCI está formada por un número relativamente pequeño de células (aproximadamente 50), la proliferación celular incrementa rápidamente este número, sin que apenas se produzca ya reducción en el volumen celular (característica que ha marcado las divisiones anteriores durante el proceso de segmentación). A la vez que se produce este incremento en el número de células, la MCI deja de ser una estructura sin organización aparente y por procesos complejos, aún poco conocidos, se constituye en una masa ligeramente aplanada o discoidal que recibe el nombre de disco embrionario (Figura 1). Las células de este disco comienzan a organizarse en dos láminas: una superior, ya que coincidirá con la región dorsal del futuro embrión, y que en estas primeras etapas limita la cavidad amniótica, y otra inferior, ya que limita con el saco vitelino del embrión y define la parte ventral del embrión (Figura 1A).
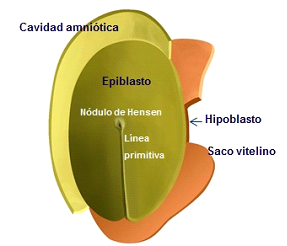
Figura 1A
Figura 1. Secuencia de dibujos que ilustran los cambios característicos del proceso morfogenético basado en la formación del embrión trilaminar a partir de la constitución del disco embrionario.
Figura 1A. Observación desde la cavidad amniótica, apreciándose las células del epiblasto constituyendo el nódulo de Hensen y la línea primitiva.
Estas dos capas reciben el nombre de epiblasto e hipoblasto y son las que constituyen en conjunto el blastodermo, y puede considerarse la primera determinación de las células que van a dar lugar al embrión propiamente dicho, ya que las células del trofoectodermo han seguido un proceso de diferenciación paralelo para dar lugar a las distintas estructuras placentarias.
El disco embrionario y más concretamente el blastodermo, es el origen de todos los tejidos embrionarios y parte de los tejidos extraembrionarios, y se ha establecido ya como una bicapa de varios cientos de células, aproximadamente en el día 10 del desarrollo embrionario (Rawles, 1943).
La formación de un nuevo organismo a partir de esta estructura necesita por una parte del establecimiento de unos ejes corporales para establecer la identidad anterior-posterior, dorsal-ventral y la del eje de simetría (izquierda-derecha), y por otra parte la formación de las distintas capas o láminas que con sus tipos celulares dan lugar a todos los órganos: ectodermo, mesodermo y endodermo (Rosenquist, 1970).
Los mecanismos mediante los cuales se establecen los ejes corporales en los vertebrados son muy complejos y posiblemente estén de alguna manera establecidos incluso desde la propia fecundación (véase artículo de Formación Continuada de Álvarez-Miguel et al., 2006). La explicación de cómo unas células del disco embrionario consiguen establecer una identidad posterior y anterior queda fuera del enfoque de este trabajo, pero en cualquier caso esta especificación de unas células del blastodermo va a determinar que se inicie el complejo proceso de migración y relocalización de células, orquestado principalmente por la línea primitiva, que se conoce como gastrulación.
3. Gastrulación: la línea primitiva y la migración celular
Uno de los procesos fundamentales en las fases iniciales del desarrollo embrionario consiste en la formación de una tercera capa embrionaria, de tal modo que el disco embrionario bilaminar, constituido por epiblasto (o ectoblasto) e hipoblasto, llegará a configurarse en un disco trilaminar, constituido por ectodermo (epiblasto en las fases previas), endodermo y mesodermo (Garcia-Martinez et al., 1993; 1997).
En fases precoces del desarrollo, las células del ectoblasto (epiblasto) inician dos procesos fundamentales, que ocurren de forma concomitante. En primer lugar, las células se dividen, proliferan, incrementan su número, lo cual conlleva al segundo aspecto, necesitan migrar, desplazarse hacia nuevas localizaciones y ocupar nuevas posiciones en el embrión. Estos procesos celulares hacen que numerosas células del epiblasto se dirijan hacia el hipoblasto, desplazando las células del mismo, para ser sustituidas por una nueva capa celular, el endodermo. En este momento, la capa de células denominada ectoblasto comienza a denominarse ectodermo (Garcia-Martinez y Schoenwolf, 1992; Schoenwolf et al., 1992; Hatada y Stern, 1994).
A partir del embrión bilaminar, constituido por ectodermo y endodermo, se inicia el proceso de gastrulación, mediante el cual se constituye la tercera capa embrionaria, el mesodermo, que se localizará entre las dos capas anteriores (Figura 1B).
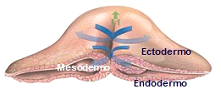
Figura 1B
Figura 1B. Sección transversal ilustrando la migración de las células del epiblasto a través del nódulo de Hensen y la línea primitiva, para ir formando progresivamente el mesodermo.
El inicio del proceso de gastrulación se caracteriza por los cambios morfogenéticos que tienen lugar en el embrión, ya que las células del ectodermo se dividen y migran, para formar la tercera capa, el mesodermo. Las células ectodérmicas, con gran capacidad de proliferación, están sometidas a diferentes corrientes de migración celular, que se identifican en dos direcciones, fundamentalmente: una corriente de células en sentido látero-medial, y una corriente de migración celular en sentido rostro-caudal. De este modo, cuando las células más laterales y las células más rostrales llegan al centro del embrión, se invaginan, a nivel de la línea media, constituyendo la denominada línea primitiva (Figura 1B, C), estructura longitudinal situada a lo largo del eje rostro-caudal del embrión, de característica fundamentalmente dinámica, lo cual indica que la línea primitiva es diferente en cada momento del desarrollo, dependiendo de las células que van ingresando a través de ella, para formar la tercera capa, el mesodermo, entre ectodermo y endodermo, constituyéndose así el embrión trilaminar (Alvarez et al., 2006).
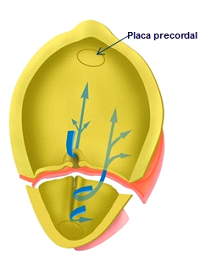
Figura 1C
Figura 1C. Embrión en fase de gastrulación, sometido a una sección transversal, que permite la observación de las corrientes de migración celular y el establecimiento de las tres capas: ectodermo, mesodermo y endodermo. Nótese en el polo cefálico la región de la placa precordal, donde el ectodermo y el endodermo están fusionados, impidiendo el paso de células de la notocorda en sentido rostral.
La línea primitiva es pues el aspecto morfológico que presentan las células cuando están ingresando a través de ésta para formar el mesodermo. Las células que siguen una migración rostro-caudal se invaginan a nivel de la zona más rostral de la línea primitiva, y forman sucesivamente, a lo largo de la capa media del embrión, una estructura de aspecto alargado, central, como una cuerda que recorre el embrión, denominada la notocorda, o también mesodermo axial, en el eje embrionario longitudinal. Las células que siguen una migración látero-medial, al invaginarse a través de la línea primitiva, formarán, a cada lado del embrión, el mesodermo propiamente dicho.
El análisis detallado de la migración celular a través de la línea primitiva pone de manifiesto que puede distinguirse un sector, claramente definido, de significación funcional diferente, en el extremo más rostral de la línea primitiva, en la posición donde se están invaginando las células de la posición rostral del ectodermo, las células que darán lugar a la notocorda (Schoenwolf et al., 1992). Esta zona inicial, rostral, de la línea primitiva se denomina, por sus características embrionarias, el organizador, descrito en ratón en los primeros estudios embrionarios, denominándose, en honor al autor que lo describió, el nódulo de Hensen. Se trata pues también de una estructura dinámica, que se encuentra en el extremo rostral de la línea primitiva, que modifica su posición en sentido rostro-caudal a medida que van ingresando mayor número de células para ir configurando la notocorda, de rostral a caudal, en el embrión (Figura 1D).
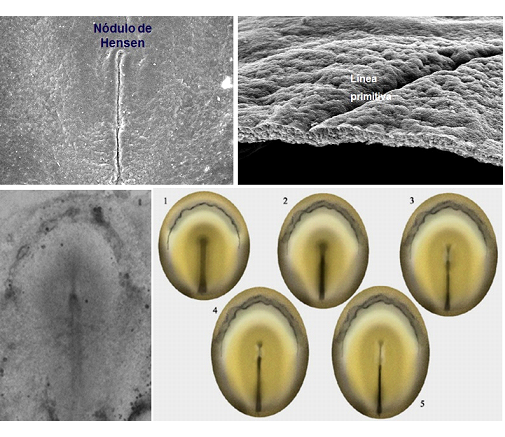
Figura 1D. La lámina ilustra el proceso de formación del nódulo de Hensen y la línea primitiva. Las imágenes superiores corresponden a microscopía electrónica de barrido. En el lado izquierdo una visión del nódulo de Hensen y la línea primitiva; a la derecha el embrión seccionado mostrando la constitución de las capas características. Las imágenes inferiores muestran a la izquierda una micrografía óptica de un embrión de ave en fase de gastrulación, ilustrado secuencialmente (del 1 al 5) en los esquemas de la imagen de la derecha.
Es interesante hacer especial énfasis en el hecho de que la notocorda, formada a partir de la línea de migración celular rostro-caudal, de las células que se invaginan a través del nódulo de Hensen, no llega a configurarse entre las capas ectodérmica y endodérmica desde las posiciones más craneales o rostrales del embrión, ya que a este nivel existe una íntima unión entre ectodermo y endodermo, donde no pueden penetrar células de la notocorda. Esta zona rostral embrionaria, donde ectodermo y endodermo están fuertemente fusionados, y no permiten la llegada de células que puedan formar notocorda se denomina la placa precordal (Figura 1C), de gran significación funcional en la constitución del extremo cefálico del eje longitudinal embrionario.
Las características dinámicas del proceso de gastrulación determinan el aspecto progresivo del desarrollo en estas fases iniciales. Dado que las células de las posiciones rostrales inician estos procesos de división y migración antes que las células de las posiciones embrionarias más caudales, el proceso del desarrollo embrionario sigue también una secuencia rostro-caudal. Un embrión mostrará ya sus tres capas embrionarias, ecto-, meso- y endo-dermo, en el sector rostral (craneal, o cefálico), mientras que el sector caudal aún estará en fase de formación de línea primitiva (Figura 1C). Es pues de gran relevancia observar que el nódulo de Hensen, y el resto de la línea primitiva, se desplazan en el embrión en sentido cráneo-caudal durante el desarrollo; el nódulo de Hensen va ocupando sucesivamente una posición más caudal, y por ello la línea primitiva va siendo progresivamente de una menor longitud (Figura 1D).
4. Destino y especificación celular durante la gastrulación
La configuración de un embrión trilaminar, constituido como consecuencia del proceso de gastrulación, aún mantiene el aspecto morfológico bidimensional, ya que dos de sus ejes (longitudinal y transversal) siguen predominando llamativamente sobre su tercer eje, determinado por el grosor, extremadamente fino, del embrión, a pesar de tener ya tres láminas (Lopez-Sanchez et al., 2001).
Es a partir de esta fase cuando el embrión comienza a crecer para ir adquiriendo progresivamente su aspecto tridimensional. Para ello el embrión mostrará dos vías fundamentales de actuación: i) cada una de las tres capas se desarrollará para formar los órganos y aparatos que específicamente se diferenciarán a partir de cada una de ellas, y ii) el cuerpo embrionario se incurvará en sentido céfalo-caudal y lateral, para ir configurando el cuerpo embrionario tridimensional. Ambos procesos tienen lugar de forma concomitante, de tal modo que a la vez que se diferencian cada una de las capas embrionarias, el cuerpo embrionario se va plegando.
Cada una de las tres láminas embrionarias seguirá patrones de diferenciación y morfogénesis específicos para cada una de ellas (Figura 2).
Aunque el desarrollo de cada capa dará lugar a distintos órganos, aparatos y sistemas, es de especial relevancia tener en cuenta que el desarrollo de cada capa es coincidente y concomitante en el tiempo con el desarrollo de las dos capas restantes. Las tres capas se van desarrollando simultáneamente. Además, durante el proceso morfogenético de cada una de ellas, existen importantes interacciones tisulares, celulares y moleculares entre los diferentes componentes de cada capa, y de los componentes de las tres capas entre sí, de tal modo que los procesos que ocurren en una determinada capa embrionaria repercuten en el desarrollo de las demás. Teniendo en cuenta estas características, plantearemos a continuación, de forma individual, el desarrollo de cada una de las tres capas (Lopez-Sanchez et al., 2005).
Ectodermo
El ectodermo determina la capa más externa (superficial) del embrión (Figura 2). Por ello, formará parte de las paredes que constituyen el espacio que rodea al embrión: el saco amniótico. En efecto, de los límites periféricos del ectodermo se diferencian un grupo de células, los amniocitos, que continuándose desde el ectodermo se disponen cerrando la cavidad, en cuyo interior queda coleccionado el líquido amniótico.
Las células ectodérmicas muestran a continuación dos zonas bien definidas: una banda longitudinal, central, desde el polo embrionario craneal hasta el polo caudal, el ectodermo neural, así denominado por contener las células que darán lugar a la formación del sistema nervioso, y el resto de la superficie ectodérmica, el ectodermo no-neural, que dará lugar fundamentalmente a la capa de células cutáneas más superficiales, la epidermis.
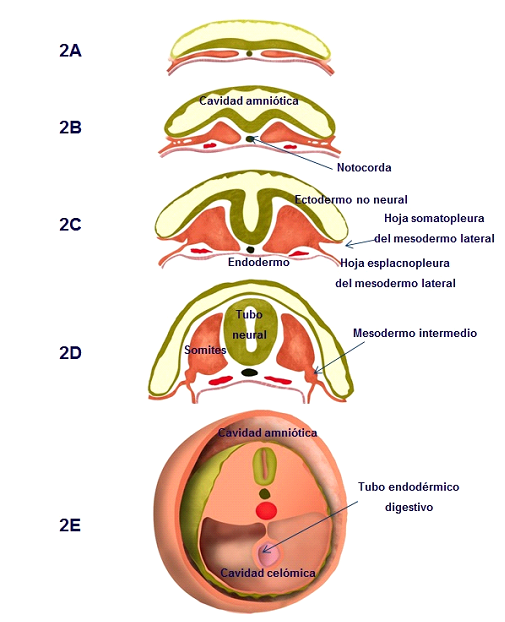
Figura 2. Secuencia de dibujos ilustrando los cambios morfogenéticos característicos de cada una de las capas embrionarias, ilustrando también el proceso de flexión lateral que da lugar al cierre del cuerpo embrionario y adquisición progresiva de su característica tridimensional.
Mesodermo
Se trata de la capa que muestra los cambios morfogenéticos más llamativos, dando lugar a un gran número de órganos y aparatos.
Aunque todas las estructuras del mesodermo se desarrollarán simultáneamente en el tiempo, analizaremos en primer lugar el componente mesodérmico situado en el eje longitudinal del embrión: la notocorda. Originada a partir de la migración rostro-caudal de las células epiblásticas más craneales del embrión, la notocorda se extenderá a lo largo de todo el embrión, a excepción de la zona más rostral, donde se lo impide la presencia de la placa precordal. La notocorda tendrá un papel fundamental en los procesos de inducción neural, sobre la capa ectodérmica suprayacente. De este modo, el ectodermo neural es la región ectodérmica longitudinal que está en íntima relación con la posición longitudinal de la notocorda. Experimentos clásicos en embriones de pollo han puesto de manifiesto que la extirpación de la notocorda conduce a una ausencia de diferenciación del ectodermo neural; y viceversa, la implantación experimental de una segunda notocorda de un embrión donante induce la diferenciación de dos regiones ectodérmicas neurales. En el curso del desarrollo la notocorda se va fragmentando y regresando, de tal modo que en el organismo adulto sólo quedan unas pequeñas reminiscencias que constituyen el núcleo pulposo de los discos intervertebrales de la columna vertebral (que con frecuencia se desplazan de su localización habitual, ocasionando las hernias discales).
El segundo componente de esta capa, el mesodermo propiamente dicho, localizado a ambos lados de la notocorda, muestra un llamativo proceso morfogenético, basado en la división, de medial a lateral, en tres diferentes sectores: el mesodermo para-axial, en relación con la notocorda, el mesodermo intermedio y el mesodermo lateral.
El mesodermo para-axial se caracteriza por su división progresiva en sentido céfalo-caudal, estableciendo la presencia de pares de acúmulos celulares localizados a cada lado de la notocorda, de este modo se constituyen los pares de somites. Cuanto mayor es el número de pares de somites, más avanzado es el estadio de desarrollo embrionario, considerándose pues este dato como uno de los principales criterios para determinar la edad del embrión. Los somites están sometidos a la diferenciación de sus células, determinándose la presencia de tres líneas de diferenciación celular: dermotomo, esclerotomo y miotomo. El dermotomo es el componente de los somites que se relacionará íntimamente con el ectodermo (formador de la epidermis) y se diferenciará para dar lugar a la dermis de la piel. El esclerotomo dará lugar a la formación de estructuras cartilaginosas y óseas, fundamentalmente las costillas y las vértebras del raquis. El miotomo es el componente celular destinado a la formación de las estructuras musculares. Es de especial relevancia el dato biológico del desarrollo basado en que estos procesos tienen lugar en íntima relación con la notocorda, por ello la columna vertebral se inicia a nivel cervical y no a nivel craneal, ya que la notocorda no alcanza los niveles más rostrales, debido a la presencia de la placa precordal, anteriormente mencionada.
El mesodermo intermedio también se denomina, globalmente, nefrotomo, ya que estos cordones longitudinales, a cada lado del embrión, estarán implicados fundamentalmente en el desarrollo de los riñones y sectores más proximales de las vías urinarias.
El tercer componente, el mesodermo lateral, se caracteriza por la formación de láminas celulares que rápidamente se dividen en dos capas, una superficial, en relación con el ectodermo, la hoja somatopleura del mesodermo lateral, y otra profunda, en relación con el endodermo, que constituye la hoja esplacnopleura del mesodermo lateral. Cada hoja de mesodermo lateral se fusionará con la del lado opuesto a nivel de la línea media del embrión, ya que éste se va plegando en sentido látero-medial formando el cuerpo embrionario. De este modo se constituye la cavidad celómica, que en el organismo adulto determinará la formación de las serosas (pericárdica, pleural y peritoneal).
El mesodermo es un buen ejemplo de la especificación del destino durante el proceso de gastrulación (Lopez-Sanchez et al., 2009). Experimentos realizados por nuestro grupo, han puesto de manifiesto, mediante trasplantes de segmentos de la línea primitiva entre embriones de pollo y codorniz, que las células que migran a través de esta estructura adquieren sus compromisos de diferenciación en función del estadio del desarrollo. Células presomíticas de la línea primitiva pueden ser destinadas a otras localizaciones si son trasplantadas a segmentos de la línea primitiva que son prospectivos de otros órganos, por ejemplo el corazón (Figura 3).
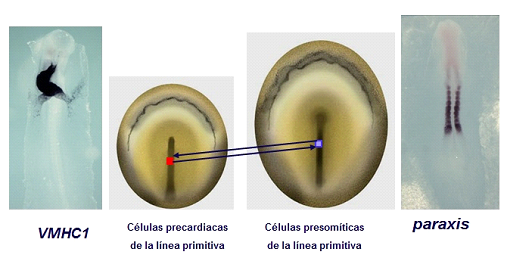
Figura 3. Procedimiento experimental mediante el cual realizamos trasplantes de sectores de la línea primitiva. El trasplante de células presomíticas de la línea primitiva a zonas precardiacas da lugar a que las células presomíticas formen corazón, y expresen el gen específico VMHC1. Y viceversa, el trasplante de células precardiacas de la línea primitiva a zonas presomíticas da lugar a que las células precardiacas formen somites, y expresen el gen específico paraxis.
Endodermo
Se trata de la capa embrionaria más profunda, en íntima relación con el saco vitelino. Es la que muestra los cambios morfogenéticos menos llamativos, adoptando una actitud aparentemente pasiva durante el desarrollo inicial, ya que se limita a seguir el proceso de incurvación embrionaria, dando lugar a la constitución del tubo endodérmico, que recorre el embrión longitudinalmente desde la boca primitiva (estomodeo) hasta el ano (membrana cloacal). Las células del endodermo constituirán fundamentalmente las estructuras del tubo digestivo, en referencia fundamentalmente a la mucosa digestiva (Figura 2E).
5. Comienzo de la organogénesis: sistema nervioso y sistema cardiovascular.
Aunque todas las estructuras referidas presentan un desarrollo simultáneo a lo largo de las diferentes fases del desarrollo inicial referido, es evidente que algunos órganos y aparatos muestran indicios de diferenciación antes que otros. Es el caso del sistema nervioso y sistema cardiovascular.
Sistema nervioso
Uno de los cambios morfogenéticos más precoces que pueden observarse a nivel del desarrollo es la diferenciación del ectodermo neural. Alrededor de la tercera semana de gestación se inicia el desarrollo del sistema nervioso. Este proceso inicial se denomina neurulación, que incluye la formación de la placa neural y los pliegues neurales, y su cierre para formar el tubo neural, aproximadamente hasta la cuarta semana del desarrollo (Figura 2). La placa neural se constituye mediante un engrosamiento del ectodermo que se relaciona en principio con el nódulo de Hensen y posteriormente con el mesodermo axial, la notocorda, prolongándose en sentido cráneo-caudal (Garcia-Martinez et al., 1993). Posteriormente, la placa neural se invagina a lo largo de su eje longitudinal para formar el canal neural y seguidamente el surco neural, con los pliegues neurales a cada lado. Los pliegues neurales se aproximan tanto que el surco neural llega a cerrarse progresivamente, constituyendo el tubo neural (Figura 2). Este proceso de cierre del tubo neural se inicia en la zona central del embrión, extendiéndose progresivamente hacia los extremos rostral y caudal, y cerrándose finalmente los orificios de los extremos, los neuroporos, craneal y caudal (Figura 4).
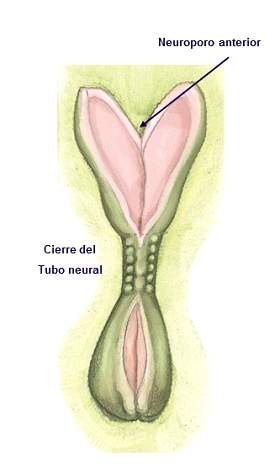
Figura 4. Visión craneal, desde el saco amniótico, mostrando la formación del tubo neural, desde la zona central del embrión hacia los polos cefálico y caudal del mismo.
El tubo neural dará origen al sistema nervioso central: encéfalo y médula espinal (Garcia-Martinez et al., 1993). Asimismo, la luz del tubo neural, el conducto neural, formará las cavidades ventriculares y el conducto ependimario, respectivamente. De la región más dorsal del tubo neural se originan las células de la cresta neural que darán lugar a numerosos tipos celulares diferenciados: 1) las células neuronales y gliales de los sistemas nerviosos sensoriales, simpático y parasimpático; 2) las células productoras de adrenalina de la glándula suprarrenal; 3) las células de la epidermis que contienen pigmentos; 4) muchos de los componentes de los tejidos conectivos y esqueléticos de la cabeza.
Cuando está finalizando el cierre del neuroporo rostral, el segmento de tubo neural más craneal (rostral al nivel del cuarto par de somites), tiene lugar el desarrollo del encéfalo. Inicialmente, este sector del tubo determina un proceso morfogenético caracterizado por la formación de las tres vesículas cerebrales primarias: prosencéfalo (cerebro anterior o rostral), mesencéfalo (cerebro medio) y rombencéfalo (cerebro caudal). Posteriormente el prosencéfalo se divide en dos vesículas, telencéfalo y diencéfalo, en tanto que el rombencéfalo se divide en metencéfalo y mielencéfalo, constituyéndose de este modo las cinco vesículas cerebrales secundarias (Figura 5).
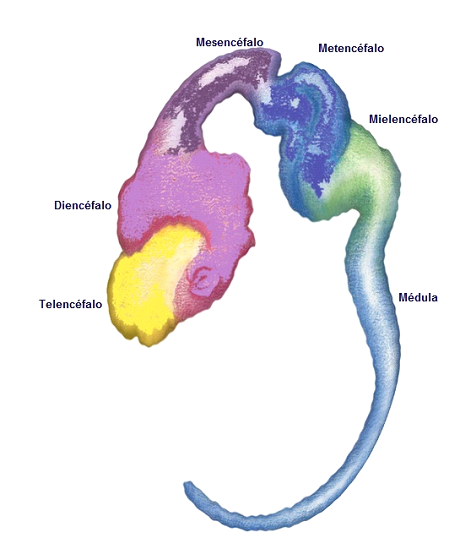
Figura 5.Establecimiento de la fase característica de cinco vesículas para la constitución del sector encefálico a nivel del sistema nervioso central.
Estructuralmente, la pared del tubo neural se compone de neuroepitelio cilíndrico pseudoestratificado, grueso, que evoluciona según se configure la estructura histológica. A nivel medular, los cuerpos o somas neuronales (sustancia gris) se localizarán en la zona central, zona ventricular, mientras que de modo gradual, la zona periférica, zona marginal, da origen a la sustancia blanca, a medida que sobre ella crecen las prolongaciones desde los somas neuronales de la médula espinal, encéfalo y ganglios raquídeos; sin embargo, a nivel encefálico, la sustancia gris adopta una disposición preferentemente superficial (corteza cerebral y cerebelosa) y la sustancia blanca una distribución más profunda.
Sistema cardiovascular
El sistema circulatorio es la primera unidad funcional en constituirse, siendo el corazón el primer órgano que comienza a funcionar (aproximadamente en la 3ª semana), para poder suministrar los requerimientos nutricionales y de oxígeno, que no pueden ser satisfechos por difusión cuando el embrión comienza a ser más complejo (Kirby, 2007). Se desarrolla inicialmente como una estructura tubular sencilla, hasta constituir un órgano maduro, tetracameral, con un alto grado de complejidad (Gonzalez-Sanchez y Bader, 1990; Harvey, 2002; Brand, 2003).
Desde el punto de vista experimental, hemos puesto de manifiesto (Garcia-Martinez y Schoenwolf, 1993; Schoenwolf y Garcia-Martinez, 1995), en embriones de aves, que las células precardiacas son identificables, incluso antes de la gastrulación, a nivel de la mitad rostral del ectodermo (epiblasto), laterales a la línea primitiva e inmediatamente caudales al nódulo de Hensen (Figura 6).
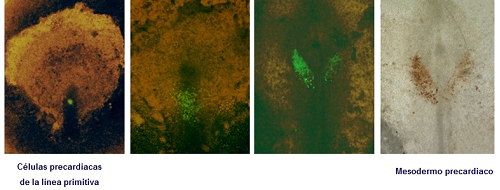
Figura 6. Embrión experimental sometido al marcaje de las células precardiacas de la línea primitiva con sustancias fluorescentes. La secuencia fotográfica pone de manifiesto la migración celular hacia el mesodermo precardiaco, detectadas posteriormente con anticuerpos frente a las células marcadas.
Al comienzo de la fase de gastrulación, las células precardiacas se dirigen hacia la línea media del embrión, se invaginan a través de zonas determinadas de la línea primitiva, situadas justo caudalmente al nódulo de Hensen, y migran rostrolateralmente en distribución bilateral, a ambos lados del nódulo de Hensen, hasta situarse a nivel de las áreas de mesodermo precardiaco, que van a contribuir a la formación del endocardio y del miocardio. Este proceso abarca en el embrión humano aproximadamente hasta el día 18 del desarrollo (Rawles, 1943; Rosenquist y DeHaan, 1966; Rosenquist, 1970). El mesodermo precardiaco establece una estrecha relación con el endodermo anterior, que ha sido implicado en el proceso de diferenciación del corazón embrionario (Stalberg y DeHaan, 1969; Fishman y Chien, 1997; Franco et al., 2002).
El mesodermo precardiaco aún indiferenciado, que se extiende a lo largo del eje antero-posterior a cada lado del embrión, sufre posteriormente un proceso de epitelialización y diferenciación que dará lugar a la formación de una estructura par, los tubos endocárdicos primitivos (Figura 7), teniendo lugar en el día 20 del desarrollo humano (Schultheiss et al., 1995).
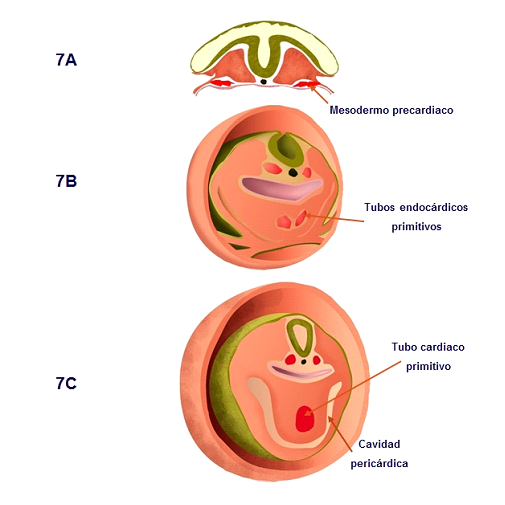
Figura 7. Secuencia que ilustra la formación de ambos tubos endocárdicos primitivos a partir del mesodermo precardiaco. Posteriormente se fusionan para constituir el tubo cardiaco primitivo, ventral a la primitiva faringe.
Debido al cierre de la placa neural y también a la elongación de las vesículas encefálicas, el sistema nervioso central crece y se expande sobre la región cardiogénica central y la futura cavidad pericárdica. En respuesta a este crecimiento del sistema nervioso y a la flexión cefalocaudal del embrión, y simultáneamente a estos eventos, los tubos endocárdicos son desplazados hacia la región cervical para posteriormente alcanzar su ubicación definitiva a nivel torácico. En este proceso los tubos se acercan y se fusionan en la línea media del embrión. La fusión comienza en el extremo cefálico de los tubos y se extiende en dirección caudal, de tal modo que se forma un tubo cardiaco único, denominado tubo cardiaco primitivo, con capacidad contráctil, y estructuralmente constituido por el endocardio, por la pared muscular o miocardio, y por el epicardio, que recubre el exterior del tubo.
En el curso del desarrollo el tubo cardiaco primitivo se incurva hacia la derecha y da lugar a la formación de una estructura denominada asa cardiaca, inicialmente con forma de “C”, y al progresar el plegamiento del cuerpo embrionario adquiere forma de “S” (Manasek et al., 1972; Lin y Taber, 1994; Levin, 1997, 2005; Levin et al., 1997; Manner, 2000; Voronov et al., 2004; Ramasubramanian et al., 2006). Este desplazamiento del tubo cardiaco primitivo constituye una de las primeras señales de asimetría izquierda-derecha en el embrión (Figura 8).
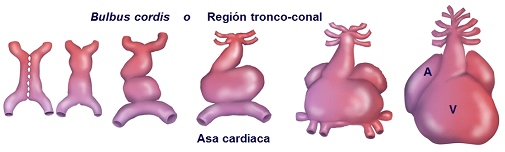
Figura 8. Proceso morfogenético que caracteriza la formación del corazón tetracameral. Desde la fusión de ambos tubos endocárdicos primitivos se establece el tubo cardiaco primitivo, que se incurva y forma el asa cardiaca. Posteriormente se divide en sectores (A: atrial, V: ventricular) y finaliza adquiriendo la forma característica del corazón adulto.
Simultáneamente a la formación del asa cardiaca se produce la segmentación o regionalización del corazón embrionario (Larry et al., 1995), de tal forma que se constituyen y diferencian las distintas partes del corazón: seno venoso en el polo caudal, atrio o aurícula y ventrículo primitivos, y bulbus cordis, región tronco-conal, o polo arterial, en el extremo cefálico. La septación o tabicación consiste en un proceso basado en la formación del septum atrial y septum ventricular, necesarios para la separación de la doble circulación del corazón, así como la formación del aparato valvular, que se localizará en la región atrioventricular y tracto de salida cardiaco (Kramer, 1942; Kirby, 2007).
En el interior del sector atrial se forma inicialmente el septum primum y posteriormente el septum secundum. El septum primum, de aspecto falciforme, desciende desde la pared superior de la aurícula primitiva, y sufre un proceso de perforación, formando el ostium primum, que será cerrado parcialmente por el desarrollo del septum secundum, a la derecha de éste. Permite el paso de sangre de la aurícula derecha a la izquierda, a través del foramen oval del septum secundum, que se cerrará totalmente en el nacimiento (Moorman y Christoffels, 2003).
El tabique o septum interventricular se forma por la organización de dos estructuras: la porción muscular y la porción membranosa. La porción muscular se origina de la capa miocárdica, en el ápice del ventrículo primitivo, permitiendo la presencia del foramen interventricular, comunicando ambos ventrículos (Escribano et al., 2006). La porción membranosa, responsable del cierre del foramen interventricular, se desarrolla a partir de las crestas tronco-conales, constituidas por células que migran desde la cresta neural, recubiertas de células de origen endocárdico (Akiyama et al., 2004).
En la división del tracto de salida cardiaco tiene lugar la separación de la arteria pulmonar y la aorta, por la presencia de las crestas troncoconales, que contribuirán además a la formación de las válvulas semilunares, aórtica y pulmonar.
6. Señalización celular
En los últimos años se ha puesto de manifiesto el papel crucial que juegan los factores moleculares en el proceso de diferenciación y morfogénesis durante el desarrollo embrionario. Numerosos trabajos previos demuestran que la expresión específica de diferentes genes es responsable de la codificación y síntesis de proteínas con capacidad de señalizar y transmitir información entre células, constituyendo las bases de la diferenciación celular. Además, es bien conocido que un determinado gen puede actuar en diferentes fases del desarrollo, y en diferentes grupos celulares. Una coordinación precisa entre moléculas inductoras, moléculas represoras y mecanismos de control de ambas dan lugar al establecimiento de rutas de señalización responsables de un determinado proceso embrionario, alcanzándose recientemente un elevado conocimiento científico de las mismas.
Los mecanismos moleculares que actúan en el desarrollo están basados fundamentalmente en dos aspectos: i) en la regulación génica por factores de transcripción, que son genes que controlan la expresión de otros genes, y ii) mediante morfógenos y señales intercelulares, mecanismo basado en la comunicación de las células a través de un receptor de la superficie celular y por la molécula (denominada ligando) que se une al mismo, permitiendo así a los receptores transmitir sus señales a procesos intracelulares.
El desarrollo del sistema nervioso constituye uno de los mejores ejemplos de la participación de la señalización molecular como base de la biología del desarrollo (Figura 9).
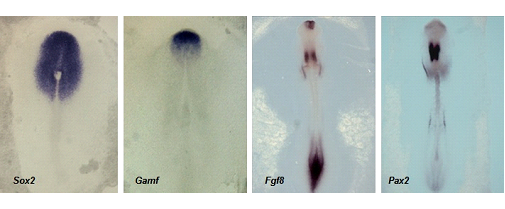
Figura 9. Colección de embriones de nuestro laboratorio, tratados mediante técnicas de hibridación in situ, que ponen de manifiesto la expresión de diferentes genes específicos, característicos de la diferenciación de las estructuras del sistema nervioso.
Una primera evidencia viene determinada por la participación de los BMPs (Bone morphogenetic proteins, proteínas morfogenéticas de hueso). BMP4: La proteína morfogenética ósea está codificada por el gen Bmp4. En ausencia de actividad de Bmp4 el ectodermo dorsal forma tejido neural por defecto. Esta proteína es inhibida por los agentes Anti-BMP (noggina, folistatina y cordina) producidos por la notocorda. Este conjunto de interacciones moleculares hace que las células ectodérmicas situadas sobre la notocorda queden comprometidas para su transformación en tejido neural, en lo que sólo representa el primer paso en la formación del sistema nervioso.
Otx2, Gbx2 y Hox son un grupo de genes que regulan la distribución regional, la subdivisión del sistema nervioso central en regiones rostro-caudales. En el desarrollo de la región del prosencéfalo y la del mesencéfalo juegan un papel determinante la expresión del gen Otx2. En el romboencéfalo estarían implicados los genes Gbx2 y Hox. La regionalización del tubo neural es particularmente evidente en la región del cerebro posterior con los genes Hox, expresados en un patrón bien definido.
La expresión de diferentes genes durante el desarrollo del corazón (Olson y Srivastava, 2006; Piedra et al., 2002) ha sido una de las principales vías de conocimiento, a nivel molecular, para la comprensión de los complejos acontecimientos que caracterizan los procesos de cardiogénesis y morfogénesis cardiaca (Figura 10).
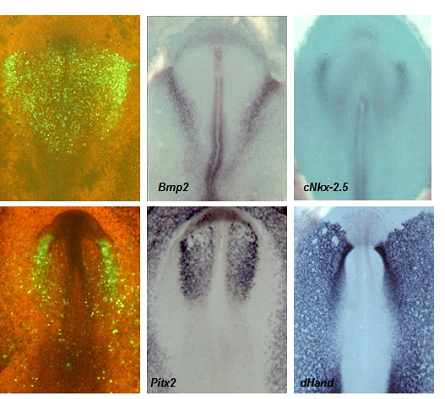
Figura 10. Colección de embriones de nuestro laboratorio, tratados mediante técnicas de hibridación in situ, que ponen de manifiesto la expresión de diferentes genes específicos, característicos de la diferenciación del corazón.
La primera manifestación de la diferenciación de células del mesodermo esplácnico hacia la formación de células cardiacas viene determinada por la expresión del factor de transcripción Nkx-2.5, como consecuencia de la inducción de miembros de las familias Fgf (Fibroblast growth factors, factores de crecimiento fibroblástico) y Bmp (Bone morphogenetic protein, proteínas morfogenéticas de hueso), que se expresan en el endodermo adyacente al mesodermo precardiaco, comprometiendo a las células de estas áreas en la vía de diferenciación de células cardiacas (Gitler et al., 2003).
En nuestro laboratorio (Lopez-Sanchez et al., 2002, 2004; Lopez-Sanchez y Garcia-Martinez, 2011) hemos puesto de manifiesto de forma experimental la relevancia de los factores de crecimiento en la inducción cardiaca. Mediante la aplicación de microesferas de heparina, cargadas con determinados factores de crecimiento, hemos demostrado la capacidad de inducción de células no precardiacas hacia la diferenciación en células específicas del corazón (Figura 11).
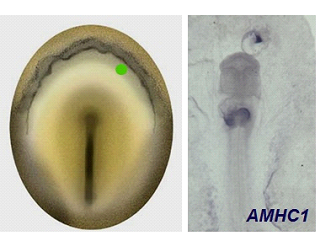
Figura 11. Embrión experimental sometido a la administración ectópica de FGF2 mediante la aplicación de una microesfera impregnada (esquema). Nótese en la micrografía del embrión experimental la expresión del gen específico de corazón, AMHC1, a nivel del corazón y a nivel del tejido ectópico inducido por FGF2.
Una de las fases fundamentales de la morfogénesis cardiaca viene definida por la formación del asa cardiaca, constituyendo la primera estructura asimétrica que aparece en el embrión. La base molecular inicial de esta diferencia izquierda-derecha vendría definida por la expresión de genes de asimetría controlados por Pitx2, y de los factores de transcripción cardiacos Nkx-2.5, Mef2, eHand y dHand (Eisenberg y Bader, 1996; Hjalt et al., 2000; Linask et al., 2002; Dong et al., 2006). La primera indicación molecular del desarrollo asimétrico del tubo cardiaco es la expresión de eHand a nivel del lado izquierdo del tubo cardiaco, mientras que dHand se expresa principalmente en el primordio del ventrículo derecho, tal como se ha puesto de manifiesto en animales de experimentación (pollo y ratón), y la eliminación experimental de estos genes origina un bloqueo de la formación del asa. En la especie humana estarían implicados (Thomas et al., 1998) los genes Hand1 (homólogo de eHand) y Hand2 (homólogo del dHand).
Recientemente han sido implicados como factores fundamentales en el desarrollo los microRNAs. Son secuencias cortas de moléculas de RNA, de 20-24 nucleótidos de longitud, que actúan como reguladores postranscripcionales de la expresión de genes. Actuarían inhibiendo la traslación del mRNA y síntesis de la proteína, o degradando el mRNA. Regulan una gran variedad de funciones del proceso de desarrollo y fisiológicas, tales como diferenciación de células madre, neurogénesis, hematopoyesis, reacción inmune o metabolismo, y han sido asociados a diversas patologías: cáncer, enfermedades autoinmunes, inflamatorias y neurodegenerativas (Fazi y Nervi, 2008; Scalbert y Bril, 2008; Sucharov et al., 2008; Thum et al., 2008; Cordes et al., 2010).
7. Significado biológico de la gastrulación
La gastrulación no es un proceso exclusivo de vertebrados y todos los animales tienen en una u otra medida una fase del desarrollo en la cual las células originadas durante las divisiones rápidas y sincrónicas del proceso de segmentación (una traducción más exacta del inglés seria partición: cleavage) adquieren una serie de características que determinan el paso de estructuras más o menos esféricas a organismos con capas, regiones, órganos y células especializadas. Este cambio se debe principalmente a que las células, una vez que adquieren un tamaño crítico (mediante reducción del gran cigoto), comienzan un proceso de migración y especificación que logra la formación de los organismos tal y como los conocemos en su fase permanente o adulta.
Este establecimiento del plan del organismo tiene en común el ingreso de células superficiales (por ejemplo el epiblasto en vertebrados) a capas más profundas, mediante desplazamientos celulares de gran envergadura para el tamaño del embrión. Desde el erizo de mar a la ballena, esta migración de células se produce a través y a la vez está coordinada por regiones específicas del embrión que actúan como centros organizadores para que, según las células pasan por estas zonas, vayan asumiendo el destino final que tendrán en el organismo, no solamente la posición que habrán de ocupar, sino también el tipo de célula que originarán.
Así, el blastoporo en invertebrados, el labio dorsal en anfibios y la línea primitiva con el nódulo en su zona más anterior actúan como verdaderos controladores del proceso de migración y consiguen que según las células pasan por estos centros, adquieran una etiqueta muy definida para la construcción final del organismo de que se trate. A modo de comparación gráfica (y ya que se trata de un artículo de formación), es como si en un reloj de arena según pasan los granos entre un compartimento y otro adquiriesen la información suficiente para formar un verdadero castillo de arena en la parte inferior.
Esta remodelación y especificación a partir de células pluripotentes (el epiblasto) para conseguir una estructura similar al organismo definitivo, y que en la especie humana ocurre aproximadamente a las dos semanas de la fecundación, es considerada también por muchos investigadores (y establecido en algunas creencias como el judaísmo) como el comienzo real de un nuevo organismo (nuevo ser), ya que es cuando la singularidad propia y no retornable se origina gracias a los complejos mecanismos de la gastrulación, que pueden llegar a ser tan únicos como el propio genoma de cada individuo. Esta apreciación, y el comentario, puede quedar fuera del enfoque de este articulo, pero podría tener un gran significado ético, y por lo tanto legislativo, que pensamos debería ser conocido por todos los embriólogos clínicos.
Agradecimientos
Agradecemos el apoyo prestado por los miembros de IERA (Instituto Extremeño de Reproducción Asistida, Badajoz) para la confección de este artículo. Agradecemos a María Pérez y Julia Anaya, ilustradoras del CCMI Jesús Usón, su trabajo en la realización de los dibujos y esquemas. Este trabajo ha sido financiado, en parte, por la Junta de Extremadura (Fondo Social Europeo), Grupos Catalogados CTS005 y CCV010.
Referencias
Álvarez-Miguel, I.S., Miguel-Lasobras, E.M., Martín-Romero, F.J., Domínguez-Arroyo, J.A., González-Carrera, E. (2006). Polaridad durante el desarrollo embrionario inicial. Rev. Asoc. Est. Biol. Rep., 11 (2): 35-45.
Akiyama, H., Chaboissier, M.C., Behringer, R.R., Rowitch, D.H., Schedl, A., Epstein, J.A., de Crombrugghe, B. (2004). Essential role of Sox9 in the pathway that controls formation of cardiac valves and septa. Proc. Natl. Acad. Sci. U S A., 101:6502-6507.
Brand, T. (2003). Heart development: molecular insights into cardiac specification and early morphogenesis. Dev. Biol., 258: 1-19.
Campbell, A., Fishel, S., Bowman, N., Duffy, S., Sedler, M., Fontes, C., Hickman, L. (2013). Modelling a risk classification of aneuploidy in human embryos using non-invasive morphokinetics Rep. BioMed. Online, 26: 477–485.
Cordes, K.R., Srivastava, D., Ivey, K.N. (2010). MicroRNAs in cardiac development. Pediatr. Cardiol., 31:349-356.
Dong, F., Sun, X., Liu, W., Ai, D., Klysik, E., Lu, M.F., Hadley, J., Antoni, L., Chen, L., Baldini, A., Francis-West, P., Martin, J.F. (2006). Pitx2 promotes development of splanchnic mesoderm-derived branchiomeric muscle. Development, 133: 4891-4899.
Eisenberg, C.A., Bader, D. (1996). Establishment of the mesodermal cell line QCE-6. A model system for cardiac cell differentiation. Circ. Res., 78: 205-216.
Escribano, D., Arbués, J., Moreno, A., López-Sánchez, C., García-Martínez, V., Galindo, A. (2006). Pallister-Killian syndrome presenting with a complex congenital heart defect and increased nuchal translucency. J. Ultrasound Med., 25: 1475-1480.
Fazi, F., Nervi, C. (2008). MicroRNA: basic mechanisms and transcriptional regulatory networks for cell fate determination. Cardiovasc. Res., 79:553-561.
Fishman, M.C., Chien, K.R. (1997). Fashioning the vertebrate heart: earliest embryonic decisions. Development, 124: 2099-2117.
Franco, D., Dominguez, J., de Castro, M., Aranega, A. (2002). Regulation of myocardial gene expression during heart development. Rev. Esp. Cardiol., 55: 167-184.
Garcia-Martinez, V., Schoenwolf, G.C. (1992). Positional control of mesoderm movement and fate during avian gastrulation and neurulation. Dev. Dynam., 193: 249-256.
Garcia-Martinez, V., Schoenwolf, G.C. (1993). Primitive-streak origin of the cardiovascular system in avian embryos. Dev. Biol., 159: 706-719.
Garcia-Martinez, V., Alvarez, I.S., Schoenwolf, G.C. (1993). Locations of the ectodermal and non-ectodermal subdivisions of the avian epiblast at stages 3 and 4 of avian gastrulation and neurulation. J. Exp. Zool., 267: 431-446.
Garcia-Martinez, V., Lopez-Sanchez, C, Darnell, D.K., Sosic, D., Olson, E.N., Schoenwolf, G.C. (1997). State of commitment of prospective neural plate and prospective mesoderm in late gastrula/early neurula stages of avian embryos. Dev. Biol., 181: 102-115.
Gitler, A.D., Lu, M.M., Jiang, Y.Q., Epstein, J.A., Gruber, P.J. (2003). Molecular markers of cardiac endocardial cushion development. Dev Dynam., 228: 643-650.
Gonzalez-Sanchez, A., Bader, D. (1990). In vitro analysis of cardiac progenitor cell differentiation. Dev. Biol., 139: 197-209.
Harvey, R. P. (2002). Patterning the vertebrate heart. Nat. Rev. Genet., 3: 544-556.
Hatada. Y, Stern, C.D. (1994). A fate map of the epiblast of the early chick embryo. Development, 120: 2879-2889.
Haunstetter, A., Izumo, S. (1998). Apoptosis: basic mechanisms and implications for cardiovascular disease. Circ. Res., 82: 1111-1129.
Hjalt, T.A., Semina, E.V., Amendt, B.A., Murray, J.C. (2000). The Pitx2 protein in mouse development. Dev. Dynam., 218: 195-200.
Kirby, M.L. (2007). Cardiac Development. Oxford University Press.
Kramer, T.C. (1942). The partitioning of the truncus and conus and the formation of the membranous portion of the interventricular septum in the human heart. Am. J. Anat., 71: 343-370.
Larry, A.T., Lin, I-E., Clark, E.B. (1995). Mechanics of cardiac looping. Dev. Dynam., 203: 42-50.
Levin, M. (1997). Left-right asymmetry in vertebrate embryogenesis. Bioessays, 19: 287-296.
Levin, M. (2005). Left-right asymmetry in embryonic development: a comprehensive review. Mech. Dev., 122: 3-25.
Levin, M., Pagan, S., Roberts, D.J., Cooke, J., Kuehn, M.R., Tabin, C.J. (1997). Left-right patterning signals and the independent regulation of different aspects of situs in the chick embryo. Dev. Biol., 189: 57-67.
Lin, I.E., Taber, L.A. (1994). Mechanical effects of looping in the embryonic chick heart. J. Biomech., 27: 311-321.
Linask, K.K., Yu, X., Chen, Y., Han, M.D. (2002). Directionality of heart looping effects of Pitx2 misexpression on flectin asymmetry and midline structures. Dev. Biol., 246: 407-417.
Lopez-Sanchez, C., Garcia-Martinez, V. (2011). Molecular determinants of cardiac specification. Cardiovasc. Res., 91: 185-195.
Lopez-Sanchez, C., Garcia-Martinez, V., Schoenwolf, G.C. (2001). Localization of cells of the prospective neural plate, heart and somites within the primitive streak and epiblast of avian embryos at intermediate primitive-streak stages. Cells Tissues Org., 169: 334-346.
Lopez-Sanchez, C., Climent, V., Schoenwolf, G.C., Alvarez, I.S., Garcia-Martinez, V. (2002). Induction of cardiogénesis by Hensen´s node and fibroblast growth factors. Cell Tissue Res., 309: 237-249.
Lopez-Sanchez, C., Garcia-Martinez, V., Lawson, A., Chapman, S.C., Schoenwolf, G.C. (2004). Rapid triple-labeling method combining in situ hybridization and double immunocytochemistry. Dev. Dynam., 230: 309-315.
Lopez-Sanchez, C., Puelles, L., Garcia-Martinez, V., Rodriguez-Gallardo, L. (2005). Morphological and molecular analysis of early developing chick require an expanded series of primitive-streak stages. J. Morphol., 264: 105-116.
Lopez-Sanchez, C., Garcia-Masa, N., Gañán, C.M., Garcia-Martinez, V. (2009). Movements and commitment of precardiac cells of the primitive streak during cardiogenesis. Int. J. Dev. Biol., 53: 1445-1455.
Manasek, F.J., Burnside, M.B., Waterman, R.E. (1972). Myocardial cell shape change as a mechanism of embryonic heart looping. Dev. Biol., 29: 349-371.
Manner, J. (2000). Cardiac looping in the chick embryo: a morphological review with special reference to terminological and biomechanical aspects of the looping process. Anat. Rec., 259:248-262.
Moorman, A.F., Christoffels, V.M. (2003). Cardiac chamber formation: development, genes, and evolution. Physiol. Rev., 83:1223-1267.
Olson, E.N., Srivastava, D. (1996). Molecular pathways controlling heart development. Science, 272: 671-676.
Piedra, M.E., Ros, M.A. (2002). BMP signaling positively regulates Nodal expression during left right specification in the chick embryo. Development, 129:3431-3440.
Ramasubramanian, A., Latacha, K.S., Benjamin, J.M., Voronov, D.A., Ravi, A., Taber, L.A. (2006). Computational model for early cardiac looping. Ann. Biomed. Eng., 34: 1655-1669.
Rawles, M. (1943). The heart forming areas of the early chick blastoderm. Physiol. Zool., 16: 22-42.
Rosenquist, G.C. (1970). Location and movements of cardiogenic cells in the chick embryo: the heart-forming portion of the primitive streak. Dev. Biol., 22: 461-475.
Rosenquist, G.C., DeHaan, R.L. (1966). In Contributions to Embryology: Migration of the precardiac cells in the chick embryo: A radioautographic study. Carn Contrib Embryol 38: 111-121.
Scalbert, E., Bril, A. (2008). Implication of microRNAs in the cardiovascular system. Curr. Opin. Pharmacol., 8:181-188.
Schoenwolf, G.C., Garcia-Martinez, V. (1995). Primitive-streak origin and state of commitment of cells of the cardiovascular system in avian and mammalian embryos. Cell. Mol. Biol. Res., 41: 233-240.
Schoenwolf, G.C., Garcia-Martinez, V., Dias, M.S. (1992). Mesoderm movement and fate during avian gastrulation and neurulation. Dev. Dynam., 193: 235-248.
Schultheiss, T.M., Xydas, S., Lassar, A.B. (1995). Induction of avian cardiac myogenesis by anterior endoderm. Development, 121: 4203-4214.
Selleck, M.A., Stern, C.D. (1991). Fate mapping and cell lineage analysis of Hensen's node in the chick embryo. Development, 112: 615-626.
Stalsberg, H., DeHaan, R.L. (1969). The precardiac areas and formation of the tubular heart in the chick embryo. Dev. Biol., 19: 128-159.
Sucharov, C., Bristow, M.R., Port, J.D. (2008). miRNA expression in the failing human heart: functional correlates. J. Mol. Cell. Cardiol., 45: 185-192.
Thomas, T., Yamagishi, H., Overbeek, P.A., Olson, E.N., Srivastava, D. (1998). The bHLH factors, dHAND and eHAND, specify pulmonary and systemic cardiac ventricles independent of left-right sidedness. Dev. Biol., 196: 228-236.
Thum, T., Catalucci, D., Bauersachs, J. (2008). MicroRNAs: novel regulators in cardiac development and disease. Cardiovasc. Res., 79: 562-570.
Voronov, D.A., Alford, P.W., Xu, G., Taber, L.A. (2004). The role of mechanical forces in dextral rotation during cardiac looping in the chick embryo. Dev. Biol., 272: 339-350.
Xiao, J., Luo, X., Lin, H., Zhang, Y., Lu, Y., Wang, N., Zhang, Y., Yang, B., Wang, Z. (2007). MicroRNA miR-133 represses HERG K+ channel expression contributing to QT prolongation in diabetic hearts. J. Biol. Chem., 282:12363-12367.
Zamir, E.A., Srinivasan, V., Perucchio, R., Taber, L.A. (2003). Mechanical asymmetry in the embryonic chick heart during looping. Ann. Biomed. Eng., 31: 1327-1336.