MUC-1 E IL6: Posibles Marcadores de Fertilidad en Reproducción Humana. Revisión
J. A. Martínez-Conejero, N. Garrido, C. Simón, A. Pellicer, J.Remohí, M. Meseguer
Departamento de Pediatría, Obstetricia y Ginecología. Facultad de Medicina de Valencia, España. Fundación IVI. Instituto Valenciano de Infertiliad. Hospital Dr. Peset.
Correspondencia: José Antonio Martínez-Conejero. Laboratorio de Andrología y Banco de Semen. Instituto Valenciano de Infertilidad. E-mail: jomarco@postal.uv.es
Con esta revisión pretendemos recopilar los datos existentes hasta la fecha en la literatura científica respecto a dos potenciales marcadores moleculares de fertilidad masculina y femenina, la interleuquina 6 (IL-6) y la mucina 1 (MUC-1), de los cuales existe un amplio conjunto de trabajos que los implican en diferentes pasos del proceso reproductivo.
Además, se han hallado ligados a ciertas situaciones patológicas que comprometen el potencial fértil, aunque no existe un consenso generalizado al respecto. A pesar de todos los esfuerzos realizados hasta la fecha, la información recopilada acerca de estas moléculas resulta insuficiente para establecer un diagnóstico y aplicar una terapia en el caso de que existiera relación con la infertilidad de causa desconocida, por lo que se requiere todavía más investigaciones al respecto.
INTRODUCCIÓN
La imposibilidad de concebir hijos o infertilidad, cada vez afecta a más parejas, y a pesar de que los avances de la ciencia permiten disponer de nuevas prácticas para solucionar tan singular inconveniente, no existe una solución definitiva para todas y cada una de las causas de infertilidad, así como tampoco se conocen todos los factores implicados que la provocan, motivo por el cual se complica más, si cabe, el tratamiento sin tener que recurrir a gametos de donantes, sobre todo en los casos más extremos.
Dichos problemas de esterilidad han sido atribuidos, tradicional y culturalmente, a la mujer. No obstante,últimamente ha venido cobrando cada vez más fuerza el factor masculino como responsable entre las posibles causas de incapacidad a la hora de tener descendencia, situándose aproximadamente en torno a un 80% o más el número de casos en los que existe alguna alteración del semen entre las parejas que acuden a un Centro de Reproducción Asistida (Garrido et al.,2001).
En los últimos años, ha ido aumentando la preocupación de la opinión pública con respecto al descenso de la fertilidad masculina. Diferentes estudios (Carlsen et al., 1992; Auger et al.,1995) basados principalmente en el examen de los análisis microscópicos de las muestras de semen según lo indicado por la Organización Mundial de la Salud (OMS) (WHO, 1999), parecen reforzar estas afirmaciones.
No obstante esta hipótesis sigue siendo polémica, y la carencia de un consenso generalizado mantiene la discusión abierta. Sin embargo en un porcentaje significativo de los varones (siguiendo los criterios de la OMS) son incapaces de embarazar a una mujer, incluso cuando la exploración de la mujer es aparentemente normal. La cuestión es que a pesar de esto, se continúa estudiando en menor proporción, haciendo mayor hincapié en las causas femeninas.
En el caso de alteración del factor masculino, existen diferentes técnicas terapéuticas capaces de mejorar las probabilidades de iniciar una gestación, como la inseminación artificial, la fecundación in vitro (FIV) o la inyección intracitoplasmática entre otras (Garrido et al., 2002), todas ellas para resolver diversas patologías seminales como oligospermia, teratospermia, astenospermia, infección en glándulas accesorias o combinaciones de ellas que derivan en la infertilidad masculina (Garrido et al., 2001). Frecuentemente la causa de infertilidad es totalmente desconocida, e individuos con un espermiograma aparentemente normal, según los parámetros de la OMS (WHO, 1999), sufren infertilidad. De ahí que sean muchos los esfuerzos por encontrar marcadores moleculares que se puedan utilizar como indicadores clínicos en circunstancias de infertilidad desconocida y para tener un mayor conocimiento de esta insuficiencia.
Los estudios básicos que conduzcan a la determinación de dichos indicadores deben pasar por una serie de etapas sucesivas. Inicialmente deben encontrarse moléculas que se hallen en el plasma seminal, en la superficie de los espermatozoides o incluso en células germinales, aunque en estos últimos, es más compleja y laboriosa la aplicación clínica. Posteriormente deben estudiarse sus posibles papeles en la fertilidad y hacer comparativas con diversos parámetros del semen y entre individuos fértiles e infértiles o con ciertas anomalias comunes en el eyaculado.
En el caso de la esterilidad femenina o factor femenino, a pesar de tener clara la influencia de distintos factores en la posibilidad de tener descendencia, como son: edad, desajustes hormonales, diferentes patologías (como por ejemplo la endometriosis) o alteraciones ovocitarias como cargas genómicas anormales o errores en alguna de las múltiples moléculas de la superficie endometrial (como la MUC-1), que pueden traer consigo problemas de implantación (Horne et al., 2001, Meseguer et al., 2001), todavía carecemos de marcadores moleculares que predigan la capacidad de una mujer de quedar embarazada.
Centrándonos en el ámbito proteico, se extrae de diversos estudios información respecto de la influencia de la expresión de ciertas moléculas, a nivel ovárico y endometrial, así como de su papel en la fisiología de la reproducción femenina. Por lo tanto, la molécula MUC-1 y otras han de ser estudiadas en detalle para poder comprender mejor la infertilidad y establecer métodos más exactos y fiables para el diagnóstico y para ofrecer con garantías de éxito, embarazos que lleguen a término sin problemas asociados.
La IL-6 y la MUC-1, son proteínas candidatas a estar implicadas, a distintos niveles, en diferentes etapas de la reproducción como la implantación embrionaria o en las propiedades fecundantes de los espermatozoides o incluso estar relacionadas entre sí. Nuestro grupo posee una amplia experiencia en el estudio de estas proteínas en diferentes aspectos de la reproducción humana.
La primera es una molécula soluble del sistema inmune, con amplio espectro de actuación y presencia en el resto de tejidos del organismo, incluido en el aparato reproductor, donde ya ha sido descrita en diferentes trabajos. Y la MUC-1 que también ha sido descrita en ambos aparatos reproductores pese a que inicialmente se caracterizó como una proteína de membrana de los diferentes epitelios.
Nuestra intención mediante esta revisión es recopilar toda la información disponible de estos dos posibles marcadores moleculares de infertilidad masculina, haciendo un repaso a aspectos bioquímicos de dichas moléculas y lo que de ellas se conoce en el campo de la reproducción.
Nuestra intención mediante esta revisión es recopilar toda la información disponible de estos dos posibles marcadores moleculares de infertilidad masculina, haciendo un repaso a aspectos bioquímicos de dichas moléculas y lo que de ellas se conoce en el campo de la reproducción.
IL-6 Estructura y Función
Las citoquinas son proteínas de bajo peso molecular, monoméricas y generalmente glicosiladas, cuya primera función caracterizada fue como molécula para la comunicación entre células del sistema inmunitario. Suelen ser sintetizadas en respuesta a patógenos o sus productos y señales relacionadas, y modulan las respuestas inflamatoria e inmunitaria. Poseen pleiotropismo, luego pueden ser sintetizadas por un amplio abanico de células, además de que presentan redundancia funcional entre ellas, es decir, pueden afectar a la actividad de otras citoquinas de forma antagónica o sinérgica, y a pesar de su corta vida media, su actividad es muy potente debido a la altísima afinidad con sus receptores(Hibi et al., 1996).
Dentro de este amplio grupo de proteínas se engloban las interleuquinas, denominadas así, porque genéricamente son producidas por leucocitos y actúan también sobre ellos. Aunque en la actualidad está ampliamente aceptado que otros tipos celulares son capaces de producirlas, y actúan sobre un vasto abanico de células, jugando un importante papel en hematopoyesis, en el hígado, en regeneración neuronal, en desarrollo embrionario y en fertilidad, entre otros aspectos (Heinrich et al., 2003).
Entre ellas encontramos a la IL-6 que originalmente fue descrita como un factor de diferenciación de células B, que inducían su maduración hasta células plasmáticas productoras de anticuerpos (Hirano et al., 1986), aunque se ha visto que también influye en placenta, hepatocitos, neuronas, células cardíacas, macrófagos y en células de la granulosa (Keck et al., 1998), etc.
Esta interleuquina es de tipo alfa: perteneciente a la superfamilia de citoquinas con cuatro hélices alfa agrupadas de cadena larga (Hibi et al., 1996). Sus funciones generales dentro del sistema inmune serían las de producir fiebre sistémica, activar expresión de moléculas de adhesión por parte del endotelio, activar a linfocitos B y T, activar expresión de proteínas de fase aguda en el hepatocito, producir la síntesis de quimioquinas que reclutan a la zona a leucocitos circulantes, inducción del desarrollo de células plasmáticas y efectos pro y anti-inflamatorios, para atajar una posible invasión de microorganismos (Van Snick et al., 1990; Barton, 1997; Tilg et al., 1997).
Estructuralmente es una proteína de 184 aminoácidos que presenta glicosilados los residuos 73 y 172, aunque previamente es traducida como una molécula de 212 aminoácidos incluyendo el péptido señal de 28 residuos (Hirano et al., 1986). Según el tipo celular presenta micro heterogeneidad en el extremo aminoterminal, por esto su peso molecular varia entre 21.5 y 28 kDa. Su gen se encuentra mapeado en 7p21-p14, se extiende unas 5 kb y presenta 4 intrones. En su promotor hay muchas secuencias reguladoras por varios estímulos (Ray et al., 1989). Los estímulos fisiológicos que desencadenan la síntesis de IL-6 son IL-1, glucocorticoides, cAMP, oncostatina M, endotoxinas bacterianas, PDGF y TNF, además de que ella puede ejercer modulación sobre sí misma según el tipo de células.
Sus funciones, descritas anteriormente, las ejerce uniéndose a su receptor (IL-6R o CD 126), que es una proteína fuertemente glicosilada de 80 kDa con 449 aminoácidos (Yamasaki et al., 1988), mapeado en 1q21 en humanos. En la parte extracelular presenta un dominio amino terminal del tipo de las inmunoglobulinas. Son generalmente varias subunidades, que a menudo están compartidas por más de un receptor, entre ellas la gp130 que estaría implicada en la transducción de la señal (Kishimoto et al., 1995). También participa de la transducción una proteína quinasa C y una adenilato ciclasa, implicadas en transducir las señales de activación al interior celular. Además ha sido descrita la existencia de un receptor soluble (siL-6R), que también interactúa con la gpl30, pero que parece estar implicado en la regulación fisiológica de la actividad de la IL-6 o bien inhibiendo la unión de ésta con el receptor de membrana o actuando como transportador de la interleuquina hasta el receptor de membrana (Kishimoto et al., 1995).
En Reproducción
Aspectos Femeninos
En los últimos años se han venido acumulando gran cantidad de evidencias que sugieren que el sistema inmune toma parte como regulador de la función reproductiva, principalmente mediante las citoquinas, que se hallan presentes en los fluidos y células implicados en reproducción.
En el tracto reproductor femenino, veíamos que se hallaba en células de placenta, en macrófagos de la zona y en células de la granulosa (Keck et al., 1998). En el caso del ovario, se ha encontrado que está asociada a funciones neoangiogénicas (Motro et al., 1990) y a otros aspectos como la regulación de la producción de progesterona observada en ratas (Gorospe et al., 1992) y actuando de forma autocrina en las células de la granulosa (Keck et al., 1998).
Además, una mala regulación de su producción podría estar afectando el ambiente folicular reduciendo la calidad del ovocito y del embrión, alterando su capacidad de implantación (Pellicer et al., 1998; Garrido et al., 2000), y por lo tanto podría estar comprometiendo la fertilidad de la mujer.
En situaciones patológicas como la endometriosis se han realizado estudios comparativos entre mujeres con y sin la enfermedad, y no se detectaban diferencias significativas ni en cantidad de la proteína ni de su mRNA en las células de la granulosa luteinizadas obtenidas de ciclos de reproducción asistida, estudiada con diferentes técnicas (Garrido et al., 2001). En estudios preliminares de nuestro grupo donde se estudiaban mujeres con y sin la enfermedad sometidas a FIV, se veían claras diferencias en la concentración de la citoquina entre líquido folicular y suero de las pacientes (Pellicer et al.,1999), siendo mayor en las pacientes con endometriosis; sin embargo en otros estudios posteriores se mostraba una disconformidad con lo anterior, es decir no hallaron diferencias, debido a la forma de purificar las células de la granulosa (Garrido et al., 2001), quedando posiblemente en el trabajo previo macrófagos y células blancas del ovario, que contribuían a la producción de IL-6.
En otra situación patológica en la que también ha sido estudiada ha sido en el síndrome de hiperestimulación ovárica (SHO), viendo que hormonas, como la hCG, producen la liberación de VEGF y al cabo del tiempo de IL-6, pudiendo estar ambos implicados en la patofisiología del SHO, ya que la IL-6 tiene propiedades vaso-activas, se encuentra en células de la granulosa y en líquido folicular, y está aumentada en mujeres que desarrollan el SHO (Albert et al., 2002).
Además, en pacientes de edad más avanzada (mayores de 37 años) el estado endocrino, paracrino y autocrino es diferente que en las jóvenes, sugiriendo que las citoquinas, especialmente las IL-6, podría estar implicada en los cambios observados durante la senescencia ovárica (Pellicer et al., 1999).
Aspectos masculinos
En cuanto a su presencia en testículo y tracto genital masculino se ha descrito la presencia o síntesis de IL-6 y/o su receptor por parte de prácticamente todas las células:Leydig, Sertoli, glándulas accesorias, células del epitelio seminífero (Kon togeorgos et al., 2002), células germinales y como veremos más adelante, en los propios espermatozoides también se producen (Huleihel et al., 2000a).
En este entorno, las citoquinas están implicadas en la regulación de células de los compartimentos tubular y endocrino (Schlatt et al., 1997). La función natural de la IL-6 podría ser la de regular y controlar la función de las células del túbulo seminífero. De hecho, en todo el túbulo seminífero existe una intricada regulación y control de la función por parte de dicha proteína (Jegou et al., 2000). Si bien, la relevancia de su síntesis y de la del receptor no han sido descritas en cuanto a la espermatogénesis, funcionalidad motora del espermatozoide o capacidad fecundante (Eggert Kruse et al., 2001).
En cuanto a su papel en fertilidad, parece haber relación entre infección en glándulas accesorias y descenso del número, concentración y movilidad de los espermatozoides. Dicho descenso parece ser atribuible a la presencia de lipo-polisacárido (LPS), que traería consigo un aumento de diversas citoquinas (IL-6 entre ellas), que inhibirían por su parte la esteroidogénesis en las células de Leydig (con IL-6 no demostrado), motivo por el cual se reduciría la producción de testosterona (Dimier et al. 2003). Por otro lado, se sabe que los andrógenos son necesarios para la correcta función reproductiva del testículo (Sharpe et al., 1994; Hales et al., 1999), el mantenimiento de la espermatogénesis (Sharpe et al., 1994; de Krester et al.,1998) y el desarrollo funcional de los espermatozoides (Chen et al., 1994; Singh et al., 1995). Se provocaría pues un descenso en la espermatogénesis y en los espermatozoides funcionales, como ilustran Diemier et al. (2003) en su revisión. En conclusión, estos polipéptidos están directamente implicados en la función testicular, bien como reguladores de la producción hormonal, bien por acción directa sobre la espermatogénesis (Rutanen et al., 1993).
Por lo tanto, en presencia de LPS, y esto viene a ocurrir cuando hay infección por bacterias Gram negativas, se produciría un aumento de IL-6 como ya hemos visto. Es decir, cuando hay infección de las glándulas accesorias, se produciría el incremento de la interleuquina y de los efectos asociados a ella, los ya mencionados, y además, se ha visto que IL-6 es un regulador paracrino específico de etapa, que actúa sobre el epitelio seminífero inhibiendo la síntesis de DNA meiótico en el preleptoteno de espermatocitos y en espermatogonias avanzadas (Hakovirta et al., 1995).
En Semen
Plasma seminal
Las interleuquinas en semen son producidas por las células del sistema inmune presentes en el eyaculado, principalmente macrófagos cuando existe una infección. Podría darse, entonces, el caso de que dicho aumento en la producción de IL-6 por causa de la infección, provocase infertilidad masculina por el mero hecho de la acumulación de ésta y otras citoquinas que perjudicarían el potencial fértil. Este efecto que podría ser producido por efectos auto o paracrino sobre diferentes células encargadas de controlar la espermatogénesis, o sobre los propios espermatozoides (Figura 1). 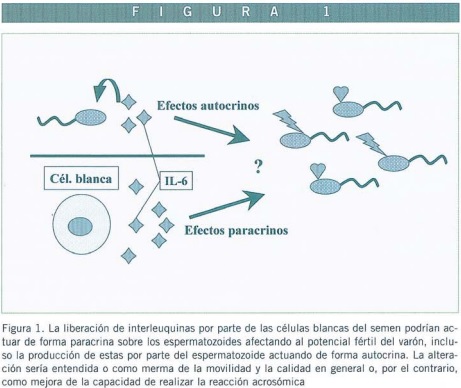
La gran mayoría de estudios realizados hasta la fecha, se han dedicado a comparar los niveles de ésta y otras citoquinas en el plasma seminal, entre individuos fértiles e infértiles y con diversos parámetros relacionados con la calidad seminal (concentración, movilidad y morfología principalmente) propuestos por la Organización Mundial de la Salud (WHO, 1999). Dichos estudios han ido encaminados a dilucidar si estas citoquinas se podrían usar como marcadores de infertilidad en el análisis de rutina del semen.
Sin embargo, el consenso entre resultados no existe, debido a los tipos de estudios realizados. La IL-6 ha sido correlacionada significativamente con algunos parámetros del semen como velocidad y linealidad de las carreras espermáticas y en ensayos de penetración espermática (Naz and Kaplan, 1994; Gruschwitz et al., 1996); también se ha encontrado una correlación positiva con el número de leucocitos en estudios de leucospermias (Shimoya et al., 1994; Sikorski et al., 2001; Egger-Kruse et al., 2001), y además del receptor (sIL-6R) con el grado de infección de las glándulas accesorias (Matalliotakis et al., 2000), con la peroxidación lipídica que sufren los espermatozoides y que podrían afectar a su funcionalidad (Camejo et al., 2001) o con proteínas del complemento como C3 (Egger-Kruse et al.,2001). Estando únicamente de acuerdo en los mayores niveles de IL-6 en los casos de infertilidad.
Otros son incapaces de confirmar dicha correlación con los parámetros habituales (morfología, concentración y movilidad) (Huleihel et al., 1996; Matalliotakis et al., 1998), ni relación con la fructosa del plasma, ni con la cantidad de anticuerpos (Egger-Kruse et al., 2001), o simplemente no aportarían la suficiente información para ser empleados como marcadores (Dousset et al.,1997). Incluso en estudios de su receptor tampoco se observaba diferencia (Huleihel et al., 1999) entre individuos fértiles y/o afectados por infertilidad. Pero sí han sido documentados altos niveles de esta citoquina en individuos con infección en las glándulas accesorias, observándose en dicha relación un alto grado de significatividad.
El estudio más completo al respecto es el realizado por Eggert-Kruse et al. en 2001, donde hace una comparativa de los distintos parámetros del semen y su calidad con las cantidades medidas de IL-6 e IL-8. Los pacientes infértiles no presentaban síntomas de infección en el tracto genital y se excluyeron a los azoospérmicos. Los resultados fueron: correlación entre ambas citoquinas pero la IL-6 en niveles más bajos; correlación positiva y significativa (P<0,05) con el pH, con número de leucocitos en plasma seminal, con niveles de proteína del complemento C3; ausencia de correlación (P>0,05) con disminución de movilidad de los espermatozoides y con test postcoital, y sin correlación con cantidad, morfología o viabilidad de los espermatozoides; ni con cantidad de fructosa en el plasma; pruebas de moco cervical de donante o pareja propia; ni con infecciones genitales, estrés, edad. Falla además al intentarlo correlacionar con la cantidad de receptor solúble (sIL-6R) (Eggert-Kruse et al., 2001).
En espermatozoides
Otros estudios han demostrado la capacidad de los espermatozoides para producir proteínas e incluso secretarlas, como exponen Huleihel et al. (2000b), con la citoquina IL-1, que los espermatozoides generan de forma constitutiva. En diclo estudio no se demuestran diferencias en cuanto a la producción entre individuos fértiles e individuos con oligoteratoastenospermia y ni siquiera se produce una inducción mediada por LPS, por lo que asumen además que no quedan leucocitos en el cultivo después del swim-up. Lo que sí son capaces de ver es que las proteínas producidas poseen bioactividad y que son liberadas al medio. Presuponiendo que dichas proteínas podrían poseer efectos de tipo paracrino sobre células de Sertoli e influir en espermatogénesis y efectos de tipo autocrino estimulando la síntesis de DNA en las fases meiótica y mitótica de las células espermatogénicas (Huleihel et al., 2000).
Siguiendo el mismo criterio, el espermatozoide podría producir también otras proteínas, entre ellas la IL-6. De hecho, otro estudio del mismo autor (Huleihel et al., 2000a) indica que al igual que con la IL-1, la IL-6 es secretada por parte de los espermatozoides y comparando entre individuos fértiles e individuos infértiles con oligoastenoteratospermia no se muestran diferencias en los niveles basales de expresión, es decir poseen expresión constitutiva y no hay diferencias entre grupos. Sin embargo, al contrario que con la IL-1, aquí sí se encuentra un incremento en la producción cuando en el medio se añade LPS, lo que implicaría que cuando aparece una infección se eleva el nivel de esta citoquina, pudiendo tener efectos paracrinos para atajar la infección, y además podría estar implicada en la fisiología y patofisiología de las funciones espermáticas, pudiendo incluso alterar la fertilidad masculina.
De hecho, la adición exógena de IL-6 a los espermatozoides mejora la capacidad de realizar la reacción acrosómica y de fecundar (Naz and Kaplan., 1994).
MUC-1 Estructura y función
Las mucinas son una familia de proteínas de elevado peso molecular (250-500 kDa) que están altamente glicosiladas y que se encuentran presentes en la superficie de los epitelíos formando el glicocálix, aunque en ocasiones se encuentran en células no epiteliales como células tumorales y células germinales. Poseen una estructura no globular, por sus abundantes residuos de prolina, alanina y glicina; y está altamente o-glicosilada, sobre todo en sus cuantiosos residuos de treonina y serina. Los oligosacáridos que se le unen se componen de N-acetil glucosamina, N-acetil galactosamina, galactosa, fructosa y ácido siálico (Devine and MacKenzie, 1992). Contienen un importante número de repeticiones en tándem que varían en número y la longitud de la unidad de repetición según la mucina, aunque también existen diferencias entre individuos en el número de repeticiones.
MUC-1 en concreto, está en el cromosoma 1 (lq21) (Gendler et al.,1990a), posee 7 exones, pero su tamaño varía de 4 a 7 kb dependiendo del número de repeticiones en tándem que posea el exón 2.
Esta proteína consta de tres partes bien diferenciadas: el amino terminal que posee una secuencia señal hidrofóbica de 13 aminoácidos, detrás de otra de 7 y delante de repeticiones en tándem degeneradas; una región central formada por las repeticiones en tándem de aminoácidos bien conservados (20 aminoácidos de 21a 125 veces) y la región carboxilo terminal que posee repeticiones en tándem degeneradas, la secuencia transmembrana y la cola citoplasmática, que puede estar fosforilada y que se asocia al citoesqueleto (Gendler et al., 1990a; Lan et al.,1990; Ligtenberg et al., 1990 y 1992; Aplin et al., 1994).
Es una proteína importantemente o-glicosilada, sobre todo en las repeticiones en tándem, ya que posee gran cantidad de serinas y treoninas. Dichas cadenas de azúcares proceden del aparato de Golgi y podrían representar hasta un 50% del peso total (Gendler et al.,199Gb).
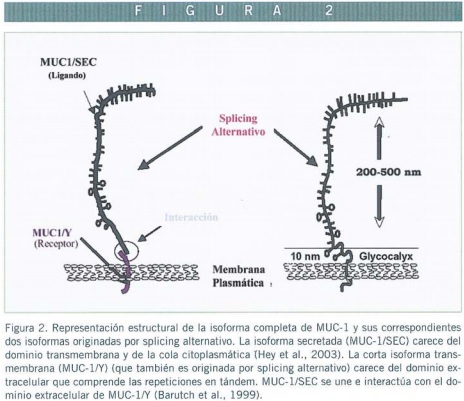 Además de todo esto, encontramos distintas isoformas de la mucina MUC-1, que se pueden explicar por dos mecanismos diferentes: el primero sería por la existencia de un sitio de corte para alguna proteasa, en una región extracelular próxima a la membrana. Este corte se produciría en el retículo endoplasmático, pero no se produce proteína secretada porque se establecerían uniones no covalentes entre ambos fragmentos que impedirían la secreción como forma libre (Ligtenberg et al., 1992; Hilkens and Boer,1998; Parry et al., 2001). Esta es la forma mayoritaria, y las otras isoformas se producirían por splicing alternativo, entre intrón 2 y exón 3, que podría sufrir el mRNA y que generaría dos péptidos, uno que sería secretado (MUC-1/SEC) (Hey et al., 2003), por carecer del dominio transmembrana, que actuaría como ligando, y otro que quedaría en la membrana (MUC-1/Y), que carecería de las repeticiones en tándem y del amino terminal (Zrihan Licht et al., 1994), actuando como receptor de membrana del anterior (Barutch et al., 1999).
Además de todo esto, encontramos distintas isoformas de la mucina MUC-1, que se pueden explicar por dos mecanismos diferentes: el primero sería por la existencia de un sitio de corte para alguna proteasa, en una región extracelular próxima a la membrana. Este corte se produciría en el retículo endoplasmático, pero no se produce proteína secretada porque se establecerían uniones no covalentes entre ambos fragmentos que impedirían la secreción como forma libre (Ligtenberg et al., 1992; Hilkens and Boer,1998; Parry et al., 2001). Esta es la forma mayoritaria, y las otras isoformas se producirían por splicing alternativo, entre intrón 2 y exón 3, que podría sufrir el mRNA y que generaría dos péptidos, uno que sería secretado (MUC-1/SEC) (Hey et al., 2003), por carecer del dominio transmembrana, que actuaría como ligando, y otro que quedaría en la membrana (MUC-1/Y), que carecería de las repeticiones en tándem y del amino terminal (Zrihan Licht et al., 1994), actuando como receptor de membrana del anterior (Barutch et al., 1999).
Dicha mucina sobresale unos 200-500 nm de la süperficie de las células, mientras que las proteínas típicas del glicocálix apenas alcanzan los 10 nm que parece indicar que juegan un papel en fenómenos de antiadhesión entre glicoproteínas de células próximas (Hilkens et al., 1992). Haciendo referencia a este hecho se ha visto, a través de deleciones, que las mucinas han de sobresalir de la superficie celular al menos entre 50 y 200 nm para ejercer su función correctamente (Wesseling et al., 1996; Komatsu et al., 1997) (Figura 2).
Las funciones que se les ha atribuido son: lubricación de mucosas y acumulación de agua para la hidratación de los tejidos, protección de las células frente a proteolisis y ataques o colonización microbiana, inhibición de funciones de células inmunes, además de posibles marcadores en algunos tipos de tumores.
Cabe destacar además, la gran capacidad antigénica que posee dicha proteína, debida sin duda al alto grado y diversidad en la glicosilación, y aunque esto a priori facilitaría su estudio por la existencia de gran cantidad de anticuerpos, la realidad es que no existen anticuerpos capaces de reconocer el amplio espectro de variantes existentes entre individuos, por lo tanto, en la práctica es complejo el estudio de esta proteína.
En Reproducción
Aspectos femeninos
La MUC-1 ha sido descrita en ambos aparatos reproductores, mientras que es considerablemente mayor el número de estudios que se han dedicado a su estudio en el tracto femenino como muestra la revisión de Meseguer et al. (1998), en la que hacen un repaso a todo el ciclo menstrual y a la etapa de implantación embrionaria en relación con la MUC-1; en el masculino ha quedado relegado a un segundo plano. Además, ha sido descrita en el embrión humano (Meseguer et al., 2001).
Como ya hemos visto, una de las funciones de las mucinas en general y de MUC-1 en particular sería la de evitar los fenómenos de adhesión fuera de lugar (Meseguer et al., 2000) debido a su longitud y rigidez, a su carencia de cisteínas y a la presencia de ácido siálico que confiere cargas negativas y la consiguiente repulsión eléctrica entre mucinas (Hilkens et al., 1992; Aplin et al., 1994).
Además de todo esto, dicha proteína evitaría la interacción entre el embrión y el epitelio materno en el proceso de la implantación, debido a la imposibilidad estérica, ejercida por la MUC-1, de encontrarse las proteínas de unión del endometrio con las del embrión. En ratones, ha sido descrita la desaparición de la MUC-1 en el endometrio en una ventana del ciclo (Aplin et al., 1997) permitiendo la implantación, sin embargo, esto sólo no sería suficiente para explicarla (Paria et al.,1993; Surveyor et al.,1995). No obstante, en humanos no se da dicha reducción durante esa ventana (Meseguer et al., 2000), lo cual representa una contradicción, aunque podría ser que gracias a otras proteínas, tipo lectinas o cadherinas, se estableciesen interacciones y así se facilitase la implantación (Meseguer et al., 1999). El papel que cabría atribuir a la MUC-1 sería el de evitar en la medida de lo posible la implantación de los embriones, dificultando en mayor grado a los que posean algún tipo de anomalía, y evitar el desarrollo de embriones anormales y el consumo de tiempo y energía que ello conlleva. Por lo tanto, esta barrera jugaría un papel de selección natural entre los posibles embriones (Hustin et al.,1990; Aplin et al., 2002).
Los embriones, asimismo provocarían durante la fase de adhesión al endometrio una reducción en la expresión de MUC-1, además de un corte proteolítico de las ya existentes, en el epitelio endometrial, acción que ejercería de forma paracrina y muy localizada en el sitio exacto de implantación. Asimismo, el propio blastocisto actuaría de forma autocrina provocándose dicho efecto, para facilitar la implantación por reducción de dicha barrera antiadhesión (Meseguer et al., 2001).
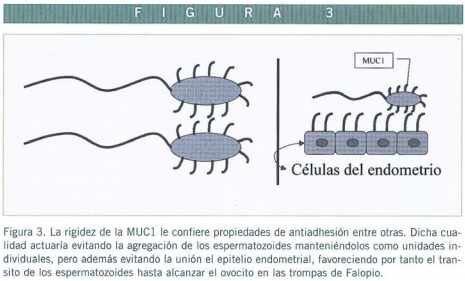
Por otro lado, la expresión de MUC-1 y su estado de glicosilación, parece jugar un papel en el tránsito de ovocitos y de espermatozoides por el tracto genital femenino (Lagow et al., 1999) (Figura 3). De hecho en el estudio de Eriksen et al. (1998), llegan a la conclusión de que gracias a las mucinas del moco cervical del tracto reproductor femenino, aumenta significativamente el porcentaje de espermatozoides móviles progresivos, principalmente traducido como un aumento de la linealidad de las carreras y de la velocidad, y además se ve reducido el porcentaje de hiperactivación (Eriksen et al., 1998).
Otra función ya mencionada es la protección frente a microorganismos. En ratones en los que se ha eliminado la MUC-1 del tracto reproductivo, que es donde normalmente quedan atrapados los microorganismos, se ha visto que se incrementa el número de infecciones, aunque se reducen las infecciones dependientes de mucinas, como Micoplasma sp. o virus común de los roedores (DeSouza et al.,1999).
Y no solamente la MUC-1 como tal ha sido descrita, sino que las diferentes isoformas se ha visto que forman parte de las trompas de Falopio y del endometrio en general, donde se expresa la forma de membrana y la secretada en sus diferentes alternativas (Meseguer et al., 2000; Hey et al.,2003).
Aspectos masculinos
En el tracto masculino, poco es lo estudiado hasta el momento sobre expresión de esta mucina pero parece ser que su presencia en diversas células germinales podría contribuir a aumentar el espacio entre células de Sertoli y células germinales, facilitando la separación de células anormales o retrasadas en el proceso de maduración. Además, podría servir como un seguro para que las células germinales madurasen en su momento exacto, protegiendo el epitelio germinal de defectos meióticos ocasionales y de retenciones de células mal diferenciadas (Franke et al., 2001). Incluso, se tienen datos de su expresión en la próstata (Ho et al., 1993), en el epidídimo (Zotter et al., 1988) e incluso en células germinales donde podemos comparar y ver la espermatogénesis (Franke et al., 2001).
Franke et al. (2001), en lo que podría ser una relación directa con fertilidad o infertilidad, detectan por diferentes técnicas la presencia de dos alelos de la proteína en el testículo adulto, sólo en espermatocitos en fase de paquiteno, en espermatocitos en fase de arresto y espermátidas, y no en células de Leydig. No se encontraron diferencias en el patrón de expresión entre individuos con espermatogénesis normal o alterada, pero la variante reconocida por el anticuerpo HMFG1 aparecía en mayor proporción en espermatocitos arrestados, o sometidos a radiación o en tubos seminíferos que estaban sometidos a degeneración por la proximidad de tumores en las células de la línea germinal, por lo que esta variante podría dar una idea de alteraciones en la espermatogénesis, ya que los espermatocitos son más frecuente e intensamente marcados con HMFG1en túbulos con espermatogénesis alterada, debido a la mayor frecuencia de la variante de MUC-1 reconocida por ese anticuerpo.
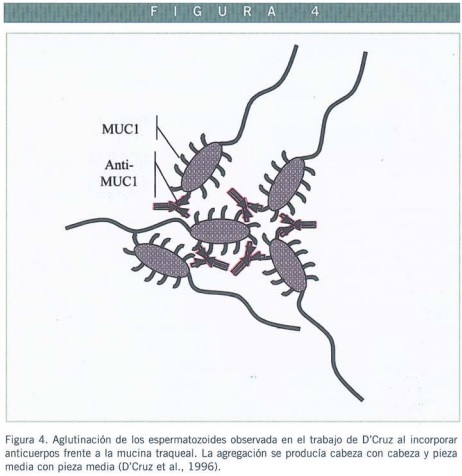
EN ESPERMATOZOIDE
Muy poco se conoce de esta proteína en el espermatozoide. Empleando una amplia batería de anticuerpos en ensayos de aglutinación de espermatozoides se encontraron resultados positivos en distintos grados de eficiencia, observando que la aglutinación de espermatozoides móviles ocurría entre cabezas, pero además entre piezas medias, formando como resultado rosetas de espermatozoides agregados (D’Cruz et al., 1996) (Figura 4).
Podría ser que la MUC-1 expresada en espermátida, contribuyese al glicocálix de la superficie de los espermatozoides, pudiendo además, estar implicada en fenómenos de antiadhesión entre ellos y con los tractos reproductores, al igual que ocurre con la MUC-8, o podría estar también implicada en la maduración y transporte de los espermatozoides y en el reconocimiento del óvulo por parte del espermatozoide (Franke et al., 2001) (Figura3).
CONCLUSIONES
Las técnicas de diagnóstico actuales no son lo suficientemente sensibles como para evaluar el problema en el varón y en consecuencia plantear una adecuada solución terapéutica o una apropiada técnica de reproducción asistida capaz de solucionar el problema. El descubrimiento de nuevas moléculas directamente relacionadas con la fertilidad masculina nos puede ayudar no sólo a mejorar el diagnóstico y el tratamiento, sino también a plantear nuevas terapias conceptivas y anticonceptivas masculinas. La existencia cada vez mayor de grupos de investigación dedicados al estudio del varón infértil nos están permitiendo comprender que el análisis de la calidad seminal no puede estar sólo basado en un estudio muy somero de la situación como es el análisis básico del semen. Los puntos desarrollados en esta revisión intentan resumir las evidencias actuales que convierten a MUC-1 y a IL-6 en marcadores de infertIlidad masculina. MUC-1 porque es una proteína de membrana que por sus características especiales podría desempeñar un papel muy importante en las interacciones del espermatozoide consigo mismo y con otros epitelios del tracto genital masculino y femenino, así como con el propio óvulo. IL-6 por ser una interleuquina pleiotrópica que como tal puede mediar en los procesos de fecundación y de interacción entre espermatozoides y otros tipos celulares del sistema inmune que podrían mediar en los procesos de transporte y quimiotactismo del propio óvulo.
AGRADECIMIENTOS
Este trabajo ha sido realizado gracias a la beca de Formación de Profesorado Universitario (FPU) concedida por el Ministerio de Educación, Cultura y Deporte a JA Martínez-Conejero y al FISss n° 0110177 concedido a los investigadores N Garrido, M Meseguer y J Remohí y al Instituto Valenciano de Infertilidad (IVI) por ofrecer sus instalaciones y equipos.
Referencias
Albert C, Garrido N, Mercader A,Rao CV, Remohí J, Simón C, et al. The role of the endothelium in the pathogenesis of ovarian hyperstimulation syndrome. Mol Hum Reprod. 2002; 8:409-18.
Aplin JD, Seif MW, Graham RA, Hey NA, Bezhad F, Campbell S. The endometrial surface and implantation. Expression of the polymorphic mucin MUC-1 and adhesion molecules during the endometrial cycle. Ann New York Acad Sci. 1994:734:103-21.
Aplin JD. Adhesion molecules in implantation. J Reprod Fertility 1997:2:84-93.
Aplin JD, Meseguer M, Simon C, Croxatto, Jones, C. MUC-1, glycans and the cell surface barrier to embryo implantation Biochemical Society Transactions. 2001;29(2):153-56.
Auger J, Kuntsman JM, Czyglick F. Jouannet P. Decline in semen quality among fertile men in Paris during the past 20 years. N Engl J Med. 1995;332:281-5.
Barton BE. IL-6: insights into novel biological activities. Clin Immunol Immunopathol 1997;85(1):16-20
Baruch A, Hartmann M, Yoeli M, Adereth Y, Greenstein S, Stadler Y, et al. The breast cancer-associated MUC-1 gene generales both a receptor and its cognate binding protein. Can Res. 1999;59:1552-61.
CameJO MI, Segnini A, Provervio F. lnterleukin-6 (IL-6) in seminal plasma of infertile men, and lipid peroxidation of their sperm. Arch Androl. 2001;47:97-101.
Carlsen E, Giwercman A, Keidmg N, Skakkebaek NE. Evidence for decreasing quality of semen during past 50 years. Br Med J.
1992; 305:609-13.
Chen H, Chandrashekar V, Zirkin BR. Can spermatogenesis be maintained quantltatively in intact adult rats with exogenously administered dihydrotestosterone? J Androl. 1994;15:132-8.
D'Cruz OJ, Dunn TS, Pichan P, Hass GG Jr, Sachdev GP. Antigenic cross-reactivity of human tracheal mucin with human sperm and trophoblasts correlates with the expression of mucin 8 gene messenger ribonucleic acid in reproductive tract tissues. Fertil Steril. 1996; 66:316-26.
de Kretser DM, Loveland KL, Meinhardt A, Simorangkir O, Wreford N. Spermatogenesis. Hum Reprod. 1998;13(Suppl. 1):1-8.
DeSouza, M.M. et al. (1999) MUC-1 Episialin: A critical barrier in the female reproductive tract. J Reprod Immunol. [in press).
Devine P y Mackenzie IFC. Mucins: structure, function and associalion with malignancy. Bioassays. 1992; 14:619-25.
Diemer T, Hales DB y Weidner W. Inmune-endocrine interactions and Leydig cell function: the role of cytokines. Andrologia.
2003; 35:55-63.
Dousset B, Hussenet F, Daudin M, Bujan L. Foliguet B y Nabet P. Seminal cytokine concentrat1ons (IL-1_, IL-2, IL-6 sR IL-2, sR IL-6), semen parameters and hormonal status in male Infertllity. Hum Reprod.1997; 12(7):1476-9.
Eggert-Kruse W, Boit R, Rohr G, Aufenanger J, Hund M y Strowitzki T. Relationsh1p of seminal plasma interleukm (IL) -8 and IL6 with semen qualily. Hum Reprod. 2001;16(3):517-28.
Eriksen GV. Carlstedt 1, Uldbjerg N, Ernst Erik. Cervical mucins affect the motility of human spermatozoa in vitro. Fertil Steril.1998; 70(2):350-4.
Franke FE, Kraus S, Eiermann C, Pauls K, Lalani E, Bergmann M. MUC-1 in normal and impaired spermatogenesis. Mol Hum Reprod. 2001; 7(6):505-12.
Garrido N, Albert C, KrOssel JS, O'Connor JE, Remohí J, Simón C et al. Expression, production, and secretion of vascular endothelial growth factor and interleukin-6 by granulosa cells is comparable in women with and without endometriosis.Fertil Steril. 2001; 76(3):568-75.
Garrido N. Meseguer M, Gám12 P, Simón C, Pellicer A, Remohí J ., Male factor and assisted reproduction techniques. ASEBIR.
2002; 7(1):10-24.
Garrido N, Meseguer M, Gil-Salom M. Pellicer A, Remohf J. Patología seminal en las parejas infértiles en un centro de reproducción asistida. X Congreso Nacional de Andrologia. Actualida Andrológica.2001; Abr28-30, Alicante, España. Libro
de Abstracts Suplemento.
Garrido N, Navarro J, Remohí J, Simón C y Pellicer A. Folicular hormonal environment and embryo quality in women with endometriosis. Hum Reprod Upd. 2000;6(1):67-74.
Gendler SJ, Cohen EP. Craston A. Duhing T, Johnstone G, Barnes D. The locus of the polymorphic epithelial mucin (PEM) tumour antigen on chromosome 1q21 shows a high frequency of alteration in primary human breast tumours. Int J Cancer.1990a;45(3):431-5.
Gendler SJ, Lancaster CA, Taylor-Papadlmitriou J, Duhig T, Peat T, Burchell J, et al. Molecular cloning and expression of human tumour associated polymorphic epithelial mucin. J Biol Chem. 1990b;265:15286-93.
Gorospe WC, Hughes FM Jr, Spangelo BL.Interleukin-6: effects on and product1on by rat granulosa cells in vitro. Endocr.1992; 130(3):1750-2.
Gruschwitz MS, Brezinschek R. Brezinschek HP. Cytokine levels in the seminal plasma of infertile males. J Androl. 1996;
17(2):158-63.
Hakovirta H. Syed V. Jegou B. Parvmen M. Function of interleukin-6 as an inhibitor of meiotic DANN synthesis in the rat seminiferous epithelium. Mol Cell Endocrinol.1995;108:193-8.
Hales DB, Diemer T, Hales KH. Role of cytokines in testicular function. Endocrine.1999; 10(3):201-17.
Heinrich PC, Behrmann 1, Haan S, Her manos HM. Muller-Newen G, Schaper F. Principies of 1L-6-type cytokine signalling and its regulation. Biochem J. 2003; [pendiente de publicar].
Hey NA, Meseguer M, Simón C. Smorodinsky NI, Wreschner DH, Ortiz ME, et al. Transmembrane and truncated (SEC) isoforms of MUC-1 in the human endometrium and Fallopian tube. Reprod Biol Endocrinol.2003; 1(1):2.
Hibi M, Nakajima K, Hirano T. IL-6 cytokine family and signal transduction: a model of the cytokine system. J Mol Med. 1996;
74(1):1-12.
Hilkens J and Boer M. Monoclonal antibodies against the non-mucin domain of MUC-1/episialin. Tumour Biol. 1998;19:67-70.
Hilkens J, Ligtenberg MJL, Vos HL y Utvinov SV. Cell membrane-associated mucins and their adhesion modulating property. Trends Biochem Sci. 1992; 17:359-63.
Hirano T. Yasukawa K, Harada H, Taga T, Watanabe Y, Matsuda T, et al. Complementary DNA for a novel human interleukin(BSF-2) that induces B lymphocytes to produce immunoglobulin. Nature. 1986;324(6092):73-6.
Ho SB, Niehans GA, Lyftogt C, Yan PS, Cherwitz DL, Gum ET, et al. Heterogeneity of mucin gene expression in normal and neo plastic tissues.Cancer Res. 1993;53:641-51.
Horne AW, White JO, Margara RA, Williams R , Winston RM, Lalani E. MUC-1: a genetic susceptibility to infertility? Lancet.
2001; 357(9265):1336-7.
Huleihel M, Lunenfeld E, Horowitz S, Levy A, Potashnik G, Glezerman M. Production of interleukin-1-like molecules by human sperm cells. Fertil Steril. 2000;73(6):1132-7.
Huleihel M, Lunenfeld E, Horowitz S, Levy A, Potashnik G, Mazor M. et al. lnvolvement of serum and lipopolysaccharide in the productionof Interleuki-1- and interleukin-6-like molecules by human sperm cells. Am J Reprod Immunol. 2000a;43(1):41-6.
Huleihel M, Lunenfeld E, Horowitz S, Levy A, Potashnik G. Mazor M, et al. Expression of IL-12, IL-10, PGE2, sll-2R and siL-6R
in seminal plasma of fertile and infertile men. Andrologia. 1999; 31(5):283-88.
Huleihel M, Lunenfeld E, Levy A, Potashnik G y Glezerman M. Distinct expression leveis of cytokines and soluble cytokine receptors in seminal plasma of fertile and infertile men. Fertil Steril. 1996;66(1):135-9.
Hustin J, Jauniaux E y Schaaps JP. Histological study of materno embryonic interface in the spontaneous abortion. Placenta.
1990; 11:477-86.
Keck C, RaJabi Z, Pfeifer K, Bettendorf H, Brandstetter T y Breckwoldt M. Expression of Interleukin-6 and interleukin-6 receptors in human granulosa lutein cells. Mol Hum Reprod. 1998; 4(11):1071-6.
Kishimoto T, Akira S, Narazaki M, Taga T. lnterleukln-6 family of cytok1nes and gp130. Blood. 1995; 86(4):1243-54.
Komatsu M, Carraway CA, Fregien NL, Carraway KL. Reversible disruption of cellmatrix and cell-cell interactions by overex pression of sialomucin complex. J Biol Chem. 1997; 272(52):33245-54.
Kontogeorgos G, Messini 1, Kyrodimou E, Giannikaki L. Zois E, Koukoulis G, et al. Endocrine. 2002; 17(2):135-40.
Lagow E, DeSouza M, DO Carson. Mammalian reproductive tract mucins. Hum Reprod Upd. 1999; 5(4):280-92.
Lan MS, Batra SK. Q1 WN, Metzgar RS, Hollingsworth MA. Cloning and sequencing of the human pancreatic tumour mucin cDNA. J Biol Chem. 1990; 265:15286-93.
Ligtenberg MJ, Kruisharr l,Buijs F, van Meijer M, Litvinov SM and Hilkens J. Cell associated episialin is a complex containing two proteins derived for a common precursor. J Biol Chem. 1992; 267:6171-7.
Ligtenberg MJ. Vos HL, Gennissen AMC, Hilkens J. Epsilin, a carcinoma-associated mucin, is generated by a polymorphic gene encoding splice vanants with alternative amino termini. J Biol Chem. 1990;265:5573-8.
Matalliolakis I, Goumenou A, Fragouli Y, Matalliotakis G, Kyriakou D. Koumantakis E. Soluble IL-6 receptor levels in the seminal plasma of infertile palienls with accessory gland infection. Arch Androl. 2000;44(3):237-42.
Matalliotakis I, Kinakou D. Fragouli 1, Sifakis S, Eliopoulos G, Koumantakis E. lnterleukin-6 in seminal plasma of fertile and infertile men. Arch Androl.1998;41(1):43-50.
Meseguer M, Aplin JD, Caballero-Campo P. O'Connor JE, Martin JC, Remohf J, et al. Human endometnal mucin MUC-1 is upregulated by progesterone and down-regulated in vitro by the human blastocyst.Biol Reprod. 2001; 64:590-601.
Meseguer M, Caballero-Campo P, O'Connor JE, Galán A, Martín JC, Remohí J, et al. Embryonic regulation of MUC-1 in human uterine epithelial cells. J Soc Gynecol Invest; 1999 Mar10-13; Atlanta, EEUU.6(Suppl. 1).
Meseguer M, Martín J, Galán A, Remohí J, Pellicer A, Simón C. MUC-1 actúa como barrera selectiva. XXIII Congreso Nacional SEF; 2000 May-Jun; vol XVII, n3.
Meseguer M, Pellicer A y Simón C. MUC-1 and endometrial receptivity. Mol Hum Reprod. 1998; 4(12):1089-98.
Meseguer M, Simón C. Wreschner OH, Hey NA, Aplin JD. The alternatively spliced MUC-1 isoform SEC is associated with the apical surface of human endometrial epithelial cells. J Soc Gynecol lnvestig 2000; Mar21-25; Chicago, USA. 7(1Suppl):211A.
Metro B, Itin A, Sachs L, Keshet E. Pattern of lnterleukin-6 express1on in vivo suggests a role for this cytokine in neoangiogenesis. Proc Natl Acad Scien USA.1990; 87:3092-6.
Naz RK, Kaplan P. Increased levels of interleukin-6 in seminal plasma of infertile men. J Androl. 1994; 15(3):220-7.
Naz RK. Kaplan P. lnterleukin-6 enhances the fertilizing capacity of human sperm by increasing capacitation and acrosome reaction. J Androl. 1994; 15(3):228-33.
Paria BC, Das SK, Andrews GK, Dey SK. Expression of the epidermal growth factor receptor gene is regulated in mouse blastocysts during delayed implantatlon. Proc Natl Acad Sci USA. 1993; 90:55-9.
Parry S. Silverman HS, McDermott K, Will1s A, Hollingsworth MA, Harns A. Identification of MUC-1 proteolytic cleavage sites in vivo. Biochem Biophys Res Commun.2001; 283:715-20.
Pellicer A. Albert C, Mercader A, Bonilla-Musoles F, Remohí J, Simón C. The follicular and endocrine environment in women with endometriosis: local and systemic cytokine production. Fertil Steril. 1998;70(3):425-31.
Pellicer A, Garrido N, Albert C, Navarro J, Remohí J y Simón C. Cytokines in older patients undergoing in vitro fertilization: The relationship to the response to controlled ovarían hyperstimulation. J Assist Reprod Genet. 1999; 16(5):247-52.
Ray A, Sassone-Corsi P, Sehgal PB. A multiple cytokine-responsive and 2nd messenger-responsive element in the enhancer of the human interleukin-6 gene: similarities with v-fos gene regulatin. Mol Cell Biol.1989; 9:5537-47.
Rutanen EM. Cytokines in reproduction. Ann Med. 1993; 25:343-7.
Schlatt S, Meinhardt A, Nieschlag E. Paracrine regulation of cellular interactions in the testis: factors in search of a function. Eur J Endocrinol. 1997; 137(2):107-17.
Sharpe RM. Regulation of spermatogenesis. In: The Physiology of Reproduction. Knobil E, Neill JD(eds). Raven Press, New York, pp. 1363-1434.
Shimoya K, Matsuzaki N, Ida N, Okada T, Taniguchi T, Sawai K, et al. Detection of monocyte chemotactic and activating factor (MCAF) and interleukin (IL-6) in human seminal plasma and effect of leukospermia on these cytokine levels. Am J Reprod lmmunol. 1995; 34(5):311-16.
Sikorski R, Kapec E, Krzeminski A, Zaleska W. Levels of proinflammatory cytokines (II-1 alpha, II-6, TNF-alpha) in the semen plasma of male partners of infertile couples. Ginekol Pol. 2001; 72(12A):1325-8.
Singh J, O'Neill C, Handelsman DJ. lnduction of spermatogenesis by androgens in gonadotropin-deficient (hpg) mice. Endocrinology. 1995; 136:5311-21.
Surveyor GA, Gendler SJ, Pemberton L, Das SK, Chakraborty I, Julian J, et al. Expression and steroid hormonal control of MUC-
1 in the mouse uterus. Endocrinology.1995; 136:3639-47.
Tilg H. Dinarello CA, Mier JW. IL-6 and APPs: anti-inflammatory and immunosuppressive mediators. Immunol Today 1997;
18(9):428-32.
Van Snick J. lnterleukin-6: an overview. Annu Rev Immunol. 1990; 8:253-78.
Wesseling J, van der Valk SW, Hilkens J. A mechanism for inhibition of E-cadherin mediated cell-cell adhesion by the mem brane-associated mucin episialin/MUC-1. Mol Biol Cell. 1996; 7:565-77
World Health Organization. WHO laboratory manual for the examination of human semen and sperm-cervical mucus interction. Cambndge: Cambndge University Press 1999, Fourth edition.
Yamasaki K, Taga T. Hirata Y, Yawata H. Kawanishi Y, Seed B, et al. Cloning and expression of the human interleukin-6 (BSF2/IFN beta 2) receptor. Science.1988; 241(4867):825-8.
Zotter S, Hagerman PC, Lossnitzer A elal. Tissue and tumour distribution of human polymorphic epithelial mucin. Cancer Rev.
1988; 11-12:55-101.
Zrihan-Licht S, Vos HL, Baruch A, ElroyStein O, Sagiv O, Keydar I, et al. characterization and molecular cloning of a novel MUCl protein, devoid of tandem repeats, expressed in human breast cancer tissue. Eur J Biochem. 1994a; 224:787-95.