NUEVOS AVANCES EN EL ESTUDIO DE LA INTEGRIDAD DEL DNA ESPERMÁTICO
Juan G. Álvarez.
Centro de Infertilidad Masculina ANDROGEN, La Coruña. Instituto Marqués, Barcelona. Harvard Medical School, Boston, MA
Resumen: La integridad del genoma paterno es de vital importancia en el inicio y mantenimiento de un embarazo a término tanto in vivo como in vitro. La presencia en el genoma del embrión de roturas en las cadenas del DNA y/o modificaciones a nivel de nucleótidos del DNA provenientes del genoma paterno, que pueden ser reparadas o no por el ovocito tras la fecundación, no es compatible con un desarrollo embrionario y fetal normal. Recientemente se ha demostrado que la fragmentación del DNA espermático es una causa potencial de infertilidad de causa desconocida. En esta revisión, los mecanismos responsables de fragmentación de DNA en espermatozoides humanos, incluyendo la apoptosis en el epitelio de los túbulos seminíferos durante el proceso de espermatogénesis, defectos en el remodelado de la cromatina durante la espermiogénesis, el daño inducido por radicales libres (ROS) durante el transporte de los espermatozoides a través del epidídimo, la activación de caspasas y endonucleasas espermáticas y el daño inducido por la quimio y radioterapia, serán discutidos. Además, los diferentes tipos de tests utilizados para determinar la fragmentación del DNA espermático y sus aplicaciones al diagnóstico y tratamiento de la infertilidad, serán descritos. Por último, se plantearán preguntas frecuentemente formuladas sobre tests de fragmentación de DNA.
Palabras clave: fragmentación de DNA, fecundación in vitro, radicales libres, estrés oxidativo, caspasas, endonucleasas, quimioterapia, radiación ionizante.
NEW ADVANCES IN THE STUDY OF SPERM NUCLEAR DNA INTEGRITY
Abstract: The integrity of the paternal genome is of paramount importance in the initiation and maintainance of a viable pregnancy in vivo and in vitro. The presence in the embryonic genome of DNA strand breaks and/or modifications at the level of DNA nucleotides coming from the paternal genome, that have not been repaired by the oocyte after fertilization, is not compatible with normal embryo and fetal development. DNA fragmentation in human spermatozoa has been recently shown to be a potential cause of unexplained infertility. In this review, the mechanisms responsible for DNA fragmentation in human sperm, including apoptosis in the seminiferous tubules epithelium, defects in chromatin remodeling during the process of spermiogenesis, and oxygen radical (ROS)-induced DNA damage during sperm migration from the seminiferous tubules to the epididymis, the activation of sperm caspases and endonucleases and damage induced by chymo and radiotherapy, are discussed. Also the different tests used to determine DNA fragmentation in human sperm and their applications to the diagnosis and treatment of infertility, are presented. Finally, answers to frequently asked questions about DNA fragmentation testing are also included.
Key words: DNA fragmentation, in vitro fertilization, oxygen radicals, oxidative stress, caspases, endonucleases, chemotherapy, ionizing radiation.
Introducción
La integridad del genoma paterno es de vital importancia en el inicio y mantenimiento de un embarazo a término tanto in vivo como in vitro. Espermatozoides con DNA dañado pueden fecundar ovocitos en metafase II con la misma eficacia que espermatozoides con DNA intacto (Aitken et al., 1998). Sin embargo, la presencia en el embrión de ciertas modificaciones a nivel de nucleótidos y/o fragmentación en las cadenas del DNA procedentes del complemento genómico paterno (que no hayan sido reparadas por el ovocito después de la fertilización), es incompatible con un desarrollo embrionario y fetal normal.
Se estima que un 25% de las causas de infertilidad masculina son de origen idiopático. La etiología de algunas de estas causas podría estar relacionada con daño de DNA en los espermatozoides. Sin embargo, este daño podría ser reparado por el ovocito después de la fecundación. Esto va a depender sobre todo (i) de la calidad citoplasmática y genómica del ovocito, y (ii) del grado de daño en las cadenas de DNA del espermatozoide que haya fecundado al ovocito. Dado que la capacidad del ovocito de reparar este tipo de daño disminuye con la edad y que, al mismo tiempo, el nivel de fragmentación de DNA en los espermatozoides aumenta, ello podría explicar, al menos en parte, la disminución significativa en la tasa de embarazo que se observa en parejas de edad avanzada. Este tipo de daño en las cadenas de DNA podría encontrarse en embriones con un complemento cromosómico normal. Es decir, la presencia de daño de DNA no reparado por encima de un umbral crítico en embriones generados tanto in vivo como in vitro, podría explicar el paro en el desarrollo embrionario que se produce tras la implantación de embriones con un cariotipo normal. Estudios recientes sugieren que este tipo de daño se expresa a partir del día 3 de desarrollo embrionario y ha sido caracterizado como late paternal effects (Tesarik et al., 2004).
Cabría puntualizar que el daño de DNA que pudiera encontrarse en el embrión no tiene por qué estar exclusivamente relacionado con el daño de DNA en el espermatozoide que haya fecundado al ovocito. Este daño podría provenir también del ovocito o de ambos. Sin embargo, dado que (i) el análisis de fragmentación de DNA en gametos es destructivo, (ii) este daño podría variar de un ovocito a otro, y (iii) el número de ovocitos es limitado, esto no nos ha permitido estudiar hasta ahora la presencia de esta patología en ovocitos. En cambio, el uso de pruebas que permitan el estudio del grado de fragmentación de DNA (especialmente cuando se trata de daño de DNA de cadena doble) en blastómeras de embriones obtenidas para Diagnóstico Genético Preimplantacional (DGP), podría aplicarse al estudio de la integridad de la cromatina embrionaria. El uso combinado de PCR , de sondas cromosómicas y de tests de fragmentación de DNA nos permitiría estudiar de forma simultánea la presencia de enfermedades monogénicas, aneuploidías y el grado de fragmentación de DNA en el embrión permitiéndonos así seleccionar los embriones de mejor calidad genómica.
Mecanismos de fragmentación de DNA en espermatozoides
¿Cómo se produce la fragmentación de DNA en los espermatozoides? El daño de DNA en los espermatozoides puede afectar tanto el DNA mitocondrial como el nuclear y puede ser inducido por cinco mecanismos principales: (i) apoptosis durante el proceso de espermatogénesis, (ii) roturas de DNA o nicks producidos durante el remodelado de la cromatina que tiene lugar durante el proceso de espermiogénesis, (iii) fragmentación de DNA a nivel post-testicular inducida por ROS y caspasas/endonucleasas durante el paso de los espermatozoides a través del epidídimo, (iv) fragmentación de DNA inducida por caspasas y endonucleasas espermáticas, y (v) fragmentación de DNA inducida por radio y quimioterapia. De estos cinco mecanismos, quizás en el que juega un papel más importante sea la fragmentación de DNA a nivel post-testicular durante el transporte de los espermatozoides a través del epidídimo. Esto viene avalado por estudios previos que demuestran que la fragmentación de DNA es más alta en espermatozoides del epidídimo (Steele et al., 1999) y eyaculados (Greco et al., 2005; Ollero et al., 2001) que en espermatozoides testiculares (Greco et al., 2005).
1. Inducción de apoptosis durante el proceso de espermatogénesis. Durante el proceso de espermatogénesis tiene lugar un screening celular importante que resulta en la inducción de apoptosis en un 50-60% de las células germinales que entran en la meiosis I. Estas células earmarked con marcadores apoptóticos tipo Fas deberían ser fagocitadas y eliminadas por la célula de Sertoli a la cual se encuentran asociadas (Billig et al., 1996; Pentikainen et al., 1999; Sakkas et al., 1999). Sin embargo, esto no siempre va a ocurrir y un porcentaje variable de estas células germinales entran en el proceso de remodelado celular de la espermiogénesis (que es el que determina la morfología espermática) apareciendo posteriormente en el eyaculado. Con relación a este proceso de screening fallido, los resultados de un estudio reciente sugieren que existe una disociación entre la calidad genómica de la célula germinal y el remodelado espermático que tiene lugar durante la espermiogénesis (Burrello et al., 2004). Es decir, una célula germinal puede tener el núcleo “pulverizado” por apoptosis o ser aneuploide y sin embargo el espermatozoide resultante tener una morfología normal. De ahí que cuando se microinyecte un espermatozoide de morfología normal, esto no nos garantiza que el genoma sea normal. Lo que si se ha constatado es que cuando existe oligospermia, la probabilidad de que un espermatozoide con morfología normal sea aneuploide es mucho mayor que si existe normozoospermia (Burrello et al., 2004). Esto probablemente esté relacionado con un bloqueo madurativo parcial asociado a alteraciones meióticas. Por último, el hecho de que un porcentaje variable de espermatozoides en el eyaculado expresan marcadores apoptóticos del tipo de Fas, fosfatidilserina, Bcl-XL, p53 (Sakkas et al., 2002) podría utilizarse para seleccionar espermatozoides no apoptóticos en muestras de semen. Un método desarrollado recientemente en esta dirección es el utilizado para separar espermatozoides apoptóticos de espermatozoides no-apoptóticos mediante el uso de las columnas de ANnexin-conjugated MicroBeads (ANMB Microbead Kit, Miltenyi Biotec, Germany). El principio en el que están basadas estas columnas es que los espermatozoides apoptóticos expresan fosfatidilserina en la hemicapa externa de la membrana y se unen a la anexina V. Al aplicar un campo magnético a las columnas, los espermatozoides unidos a la anexina V conjugada con las micropartículas magnéticas serían retenidos en la columna, mientras que los no-apoptóticos pasarían a través de la misma (Grunewald et al., 2001). Otro método, quizás más específico, sería la selección de espermatozoides apoptóticos
por técnicas de inmunoadsorción fase sólida mediante el uso de anticuerpos anti-Fas adheridos a placas de Petri.
2. Roturas de DNA durante el proceso de espermiogénesis. Alteraciones en el proceso de remodelado de la cromatina espermática durante la espermiogénesis podrían resultar en fragmentación de DNA. McPherson y Longo han postulado que la presencia de roturas en el DNA podría ser indicativa de maduración incompleta durante la espermiogénesis. Para que se produzca el empaquetamiento de la cromatina del espermatozoide, es necesaria la actividad de nucleasas endógenas que corten y liguen el DNA durante su protaminación. Estos cortes proporcionarían una liberación de estrés torsional ayudando así al empaquetamiento de la cromatina durante el desplazamiento de las histonas por las protaminas (McPherson and Longo, 1992, 1993a, 1993b).
Alteraciones en el control de este proceso podrían resultar en roturas de DNA no reparadas. Estas alteraciones se producirían antes de la espermiación.
3. Fragmentación de DNA a nivel post-testicular. Estudios recientes demuestran que espermatozoides inmaduros que producen niveles elevados de ROS pueden inducir daño de DNA en espermatozoides maduros. Este daño se produciría después de la espermiación durante la co-migración de espermatozoides maduros e inmaduros desde los túbulos seminíferos al epidídimo (Ollero et al., 2001). Dado que los espermatozoides se encuentran en íntimo contacto en el epidídimo,
y que la vida media de los ROS es del orden de nano a microsegundos, esto facilitaría el que los ROS induzcan fragmentación de DNA a nivel del epidídimo, ya sea actuando directamente sobre el DNA o bien indirectamente mediante la activación de endonucleasas y caspasas espermáticas. Esto es consistente con el hecho de que la co-centrifugación de espermatozoides inmaduros (que producen
niveles elevados de ROS) con espermatozoides maduros resulta en la inducción de fragmentación de DNA en estos últimos, ya que en estas condiciones estos espermatozoides también se encontrarían en íntimo contacto (Twigg et al., 1998). Esto también es consistente con el hecho de que la exposición in vitro de espermatozoides maduros a ROS resulta en daño significativo de DNA (Aitken et al., 1998; Lopes et al., 1998). Por otra parte, el mismo epidídimo podría también jugar un papel activo a la hora de inducir fragmentación de DNA en los espermatozoides a su paso por el mismo, ya sea producido por radicales libres como el anión superóxido (Britan et al., 2006), el radical hidroxilo o el óxido nítrico, o bien mediante la activación de caspasas y endonucleasas espermáticas por agentes tóxicos o por factores epididimarios. En el primer caso, este tipo de daño podría prevenirse mediante el uso de agentes antioxidantes, mientras que en el segundo caso este tratamiento carecería de eficacia. Esto viene avalado por los resultados recientemente publicados por Greco et al., donde el uso de
antioxidantes produjo una reducción significativa en los niveles de fragmentación de DNA (Greco et al., 2005).
Probablemente, los espermatozoides que expresen un mayor daño de DNA sean aquellos que adquieran un menor grado de crosslinking de puentes disulfuro en la cromatina espermática durante su maduración en el epidídimo. En este sentido, estudios recientes han demostrado que, en general, el grado de fragmentación de DNA en espermatozoides eyaculados es más alto que en espermatozoides testiculares (Greco et al., 2005; Steele et al., 1999) y que en espermatozoides del cuerpo y cabeza del epidídimo (que es donde se completa el proceso de crosslinking) y que la inducción de fragmentación de DNA en los espermatozoides a su paso por el epidídimo podría estar relacionada con la calidad genómica del espermatozoide. Es decir, además del mecanismo de screening de la célula de Sertoli, al que hacíamos referencia antes, existiría otro mecanismo de screening a nivel del epidídimo dirigido a eliminar espermatozoides genómicamente defectuosos (Suganuma et al., 2005).
El daño potencial de DNA que los espermatozoides pueden experimentar a su paso por el epidídimo tiene una gran trascendencia clínica, ya que en casos de niveles elevados de fragmentación de DNA en semen y fallo en dos o más ciclos de FIV/ICSI, podría recurrirse a la microinyección de espermatozoides testiculares obtenidos mediante técnica de TESA o TESE (preferiblemente TESA, dado que es menos invasiva y gozaría de una mayor aceptación por parte del paciente y del ginecólogo). Esto viene avalado por los resultados obtenidos en un estudio publicado por Greco et al., donde la microinyección de espermatozoides testiculares en pacientes con un nivel de fragmentación de DNA en semen >15%, medido por el test TUNEL, resultó en una tasa de embarazo del 44,4%, mientras que con espermatozoides eyaculados la tasa de embarazo evolutivo fue del 0% (Greco et al, 2005).
Cabe destacar que la fragmentación de DNA inducida por el radical hidroxilo resulta en la formación de 8-OH-guanina y 8-OH-2´-deoxiguanosina en un primer estadio seguido de fragmentación de DNA de cadena doble (Cui et al., 2000). Mientras que el daño de DNA del primer tipo pudiera ser reparado por el ovocito, la fecundación de un ovocito por un espermatozoide con fragmentación de DNA de cadena doble es prácticamente irreversible e incompatible con el desarrollo de un embarazo a término. Dado que valores de fragmentación de DNA en espermatozoides eyaculados >20%, medidos por TUNEL (Sergerie et al., 2005), o >30%, medidos por el test SCSA (Evenson et al., 1999), están asociados a tasas de embarazo <5%, la pregunta que surge es qué ocurre con el 80% y 70% restante de los espermatozoides. En estos casos, la probabilidad de que un espermatozoide con un DNA normal fecunde al ovocito debería de ser del 80% y 70%, respectivamente y no <5%. Sin embargo, además de que un 20% y 30% de los espermatozoides tengan roturas en la cadena de DNA, el resto de los espermatozoides podrían tener modificaciones en las bases de DNA del tipo de 8-OH-guanina y 8-OH-2´- deoxiguanosina. Por lo tanto, la probabilidad de que un espermatozoide con DNA normal fecunde al ovocito sería mucho más baja que la esperada con un valor de fragmentación de DNA del 20% o 30%, respectivamente. Es decir, además de la fragmentación de DNA determinada del 20% y 30% (daño primario), el 80% y 70% restante de los espermatozoides tendrían otro tipo de daño (daño secundario) que no miden los tests habituales de fragmentación de DNA y que, de no ser reparado, no es compatible con el desarrollo de un embarazo a término. Este concepto ha sido designado como el “efecto iceberg” (Evenson et al., 1999).
4. Activación de caspasas y endonucleasas espermáticas. La activación de caspasas y endonucleasas en espermatozoides diferenciados por factores físico-químicos puede inducir también fragmentación del DNA espermático. Estudios previos indican que la exposición de espermatozoides de ratón in vitro a 40ºC resulta en un aumento significativo en el grado de fragmentación del DNA (Sailer et al., 1997). Más recientemente, Banks et al., han demostrado la inducción de fragmentación de DNA en espermatozoides de ratón in vivo tras haber sido expuestos los testículos a una temperatura de 42ºC (Banks et al., 2005). Dado que la fragmentación de DNA se encontró en espermatozoides aislados del epidídimo una hora después del estímulo, los autores concluyeron que el daño observado tendría que haberse producido en los espermatozoides a su paso por epidídimo y que podría ser causado por radicales libres o por activación de caspasas y endonucleasas espermáticas.
5. Fragmentacion de DNA inducida por radio y quimioterapia. Tests de fragmentación de DNA
Recientemente se han introducido una serie de tests para la determinación de daño de DNA en los espermatozoides. Estos tests incluyen TUNEL (TdT-mediated-dUTP Nick-End Labelling, Gorczyca et al., 1993), COMET (Hughes et al., 1996), CMA3 (Manicardi et al., 1995), ISNT (in-situ nick translation, Tomlinson et al., 2001), DBD-FISH (DNA Breakage Detection Fluorescence In Situ Hybridization o detección de roturas en el DNA por hibridación in situ, Fernández et al., 1998, 2000), SCD (Sperm Chromatin Dispersion Test, Fernández et al., 2003) y el SCSA (Sperm Chromatin Structure Assay, Evenson et al., 1980; 1983; 1985; 1991; 1994; 1999a; 1999b).
Un aspecto importante en la determinación de la fragmentación de DNA es el relacionado con el tipo de roturas que se produce en las cadenas de DNA (cadena sencilla versus cadena doble) y la susceptibilidad a la fragmentación de DNA. Tests de fragmentación de DNA como el SCSA, SCD (Fernández et al., 2003), o COMET a pH ácido o alcalino (Singh et al., 1988; 1989) requieren un paso inicial de desnaturalización para detectar los fragmentos de DNA o roturas potenciales en la cadena de DNA. De hecho, cuando se observa daño de DNA en condiciones de pH ácido o alcalino y no en condiciones de pH neutro se habla de puntos lábiles de rotura al tratamiento ácido o alcalino. Por el contrario, el test TUNEL (Gorczyca et al., 1993), ISNT (Gorczyca et al., 1993) y el COMET a pH neutro (Singh et al., 1988), no requieren un paso previo de desnaturalización y miden roturas auténticas de DNA de cadena sencilla (ISNT, TUNEL y COMET) y cadena doble (TUNEL y COMET). Dado que el pH intracelular en el ovocito es de 7.0, roturas de DNA de cadena sencilla o la presencia de puntos lábiles de rotura en el DNA no tendrían mayores consecuencias en la formación del pronúcleo masculino, ya que a pH neutro las cadenas de DNA no se disocian y, por lo tanto, este tipo de daño sería mucho más fácil de reparar. Por lo tanto, como una primera aproximación, podrían considerarse dos tipos de tests: (i) tests que miden “daño real” de DNA tales como TUNEL, ISNT o COMET a pH neutro, y (ii) tests que miden “daño potencial” de DNA o puntos lábiles de rotura al tratamiento ácido o alcalino y susceptibilidad a la desnaturalización de DNA, tales como el SCSA, SCD, Chromomycin A3 y COMET a pH ácido o alcalino (Singh et al., 1989). Tests que miden daño real de DNA deberían tener un mayor valor predictivo que tests que miden daño potencial de DNA. Esto está avalado por el estudio publicado por Greco et al., en el que se demuestra que la microinyección de espermatozoides eyaculados con valores de TUNEL >15% resultaron en una tasa de embarazo evolutivo por ciclo del 0% mientras que la microinyección de espermatozoides testiculares con valores <6% en un segundo ciclo de ICSI en estos mismos pacientes resultó en una tasa de embarazo por ciclo del 44.4% (Greco et al., 2005).
Es importante resaltar que si bien el test TUNEL con frecuencia se utiliza para determinar apoptosis en células, la positividad de TUNEL no es siempre sinónima de apoptosis, ya que el daño de DNA inducido por el radical hidroxilo y la radiación ionizante también resulta en fragmentación de DNA de cadena doble que puede ser detectada por el test TUNEL (Negoescu et al., 1998).
El test que más se ha estudiado hasta ahora desde el punto de vista clínico es el test SCSA desarrollado por Evenson et al. (Evenson et al., 1980; 1983; 1985; 1991; 1994; 1999a; 1999b). El test SCSA mide la susceptibilidad del DNA de los espermatozoides a la desnaturalización in situ tras la exposición a un pH ácido. El DNA de espermatozoides con una cromatina normal no se desnaturaliza, mientras que si el DNA de los espermatozoides está dañado y contiene roturas en sus cadenas, se pueden alcanzar diferentes grados de desnaturalización. Para determinar el grado de desnaturalización del DNA espermático, tras el tratamiento ácido, se tiñen las células con naranja de acridina (AO). Dado que el AO es un fluorocromo metacromático, éste emite en verde cuando se intercala en cadenas de DNA dobles (DNA intacto), y en rojo si son de cadena sencilla (es decir, DNA fragmentado). El grado de daño de DNA, es decir la intensidad de la fluorescencia verde y/o roja en los espermatozoides se mide de forma cuantitativa por citometría de flujo y se expresa como DFI (DNA Fragmentation Index). Estudios previos indican que valores de DFI >27% están asociados con fallo de embarazo en TRA (Larson et al., 2000; Larson- Cook et al., 2003). Sin embargo estudios recientes ponen en tela de juicio el valor predictivo del test SCSA, ya que se han reportado casos de embarazo en técnicas de reproducción asistida con valores de DFI >27% (Gandini et al., 2004; Bungum et al., 2004).
Quizás en la mayoría de los casos el daño de cadena sencilla detectado en el DNA espermático por tests como el test SCSA no sea incompatible con una descondensación normal de la cromatina durante el proceso de fecundación del ovocito y la formación del pronúcleo masculino. En este sentido, el recién desarrollado test SCD nos puede ayudar a comprender mejor este proceso. El test SCD está basado en el hecho de que en espermatozoides con las cadenas de DNA intactas se produce un desenrollamiento de los bucles de DNA, empaquetados en la matriz nuclear, tras haber sido expuestos a un tratamiento ácido seguido de un tratamiento con agentes reductores y detergentes. Este desenrollamiento produce un halo alrededor de la matriz nuclear que se puede visualizar por microscopía de campo claro usando tinciones habituales como el Diff-Quik. Por el contrario, si una de las cadenas de DNA está dañada, no se produce el halo, observándose núcleos condensados. Sin embargo, si los espermatozoides no se exponen a este tratamiento ácido previo, se produce el desenrollamiento de los bucles de DNA, independientemente de que el DNA esté fragmentado o no. Dado que, como ya indicamos antes, el pH intracelular del ovocito es del orden de 7.0, un espermatozoide con DNA fragmentado que haya fertilizado el ovocito, podría desenrollar sus bucles de DNA permitiendo que se produzca la descondensación de la cromatina y la formación del pronúcleo masculino. Sin embargo, todavía cabría preguntarse ¿qué ocurre con ese pronúcleo masculino en el cual existen cadenas de DNA potencialmente dañadas? ¿va a afectar el desarrollo posterior del embrión? Obviamente, la respuesta a esta pregunta va a depender de varios factores
entre los que cabe destacar el número de ovocitos obtenidos, la capacidad del ovocito de reparar este daño, si existe daño real o daño potencial y de si están afectados intrones o exones.
Dado que más del 90% del genoma humano está compuesto por intrones con secuencias repetitivas que no codifican por proteínas celulares, lo más probable es que de existir daño en las cadenas de DNA, éste no afecte a hot spots o secuencias que codifican proteínas (exones) que pudieran comprometer el desarrollo normal del embrión. Sobre este tema volveremos a hablar más adelante en la sección de preguntas y respuestas.
Dadas las limitaciones de tests como el SCSA y SCD, que miden susceptibilidad del DNA a la desnaturalización in vitro, en la predicción de tasas de fecundación y embarazo en técnicas de reproducción asistida, en la actualidad se recomienda el uso de tests como TUNEL o in-situ nick translation que miden “daño real” de DNA. La presencia de daño de cadena doble en el espermatozoide podría interferir de forma significativa con la descondensación de la cromatina espermática. Esto es consistente con los resultados de estudios que demuestran la existencia de bajas tasas de fecundación asociadas a niveles de fragmentación de DNA >10% medidos por el test TUNEL (Lopes et al., 1998; Benchaib et al., 2003). Sin embargo, como cabría esperar, el test SCSA no se correlaciona con las tasas de fecundación in vitro (Larson et al., 2000; Larson- Cook et al., 2003).
Aplicaciones al diagnóstico y tratamiento de la infertilidad ¿Cuáles son las implicaciones del daño de DNA en el diagnóstico y tratamiento de la infertilidad? La fecundación de ovocitos en metafase II por espermatozoides con daño de DNA podría conducir a alteraciones en el desarrollo del embrión, fallo en la implantación, o a un aumento en la tasa de aborto (Genesca et al., 1992; Parinaud, 1993; Twigg et al., 1998; Evenson, 1999b: Carrell et al., 2003). Ovocitos maduros con los mecanismos de reparación de DNA intactos tienen la capacidad de reparar un daño moderado de DNA en los espermatozoides (Brandiff and Pedersen, 1981; Matsuda and Tobari, 1988). Sin embargo, ovocitos inmaduros u ovocitos cuyos mecanismos de reparación no funcionen de forma adecuada (p.e., mujeres de edad avanzada) o que hayan sido dañados por factores tóxicos endógenos (p.e., radicales libres) o exógenos (p.e., radiación, tóxicos ambientales) no serían capaces de reparar este daño. Esto es consistente con la observación de que la inseminación de ovocitos de mujeres de edad avanzada -que previamente habían sido inseminados in vitro en varios ciclos con espermatozoides de baja calidad de su cónyuge y cuya transferencia resultó en fallo repetido de embarazo- con espermatozoides de donante, frecuentemente resulta en un aumento significativo en la calidad embrionaria y en la tasa de embarazo (Genescá et al., 1992; Obasaju et al., 1999). Por consiguiente, los tests de fragmentación de DNA podrían potencialmente utilizarse en el diagnóstico de fallo repetido de embarazo y abortos de repetición en TRA.
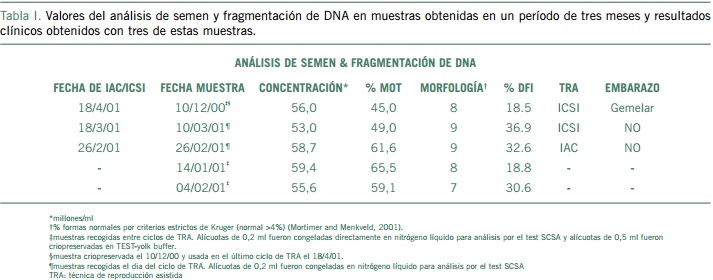
Sin embargo, no todas las muestras de semen que se obtengan de un paciente van a tener el mismo nivel de fragmentación de DNA. En un trabajo reciente, se ha encontrado que existe gran variabilidad en el grado de fragmentación de DNA en muestras de semen obtenidas de un mismo paciente en diferentes ciclos espermatogénicos (Álvarez et al., 2004). Los resultados que se muestran en la Tabla I corresponden a muestras de semen obtenidas de un mismo paciente en un período de tres meses. En esta pareja, alícuotas de las muestras de semen utilizadas en sus ciclos de reproducción asistida, fueron congeladas con TEST-yolk buffer/glicerol y alícuotas de 0,2 ml de estas mismas muestras, así como de otras tres muestras que se obtuvieron antes de empezar sus ciclos de reproducción asistida, fueron congeladas directamente en nitrógeno líquido y enviadas a un laboratorio de referencia para el análisis de fragmentación de DNA por el test SCSA. Como muestran los resultados, de las cinco muestras que se analizaron, tres tuvieron valores de DFI >27% (las tres que resultaron en fallo de embarazo) y dos <27% (una de ellas resultó en un embarazo gemelar). Dado que en esta pareja no se produjo embarazo tras dos ciclos de TRA, se decidió utilizar una de las muestras que había sido previamente criopreservada con TEST-yolk buffer/glicerol y que tenía un valor de DFI por debajo de 27%. El uso de esta muestra en un ciclo posterior de ICSI resultó en un embarazo gemelar a término.
Si bien estos resultados son todavía muy preliminares y habría que confirmarlos en un mayor número de casos, sugieren que los tests de fragmentación de DNA espermático podrían utilizarse no sólo para el diagnóstico sino también para el tratamiento de la infertilidad. Es importante destacar que a pesar de que la fragmentación de DNA varió de forma significativa en los dos ciclos espermatogénicos estudiados, los parámetros estándar de semen no mostraron una variación significativa y estuvieron dentro de los límites de la normalidad. Es decir, la causa de infertilidad de esta pareja pasaría de ser idiopática a ser de factor masculino.
Otro aspecto importante de las implicaciones clínicas del estudio de la integridad del DNA espermático está relacionado con el grado de fragmentación de DNA que se encuentra en espermatozoides testiculares vs. espermatozoides del epidídimo en oligospermias y azoospermias obstructivas (Steele et al., 2003). En este estudio, Steele et al., demostraron que el grado de fragmentación de DNA en espermatozoides testiculares de pacientes con procesos obstructivos es significativamente inferior al de espermatozoides del epidídimo obtenidos de esos mismos pacientes. Estudios previos sugieren que factores tóxicos medioambientales, radicales libres producidos por espermatozoides inmaduros, o bien derivados de un varicocele testicular o producidos por el propio epidídimo (p.e., óxido nítrico producido por iNOS epididimaria), y/o la presencia de factores proinflamatorios, p.e., IL-6, IL-8) podrían resultar en la inducción de fragmentación de DNA. Por lo tanto, en lugar de obtener espermatozoides de los túbulos epididimarios en casos de procesos obstructivos, donde se pueden acumular estos factores y también se puede producir una obstrucción iatrogénica, se recomienda obtener espermatozoides testiculares, ya sea por la técnica de TESA o TESE.
Preguntas frecuentes sobre tests de fragmentación de DNA
1. ¿A qué pacientes se les debería hacer el test?
El análisis de fragmentación del DNA espermático estaría indicado solicitarlo en los siguientes casos:
1. Fallo repetido de FIV
2. Abortos de repetición (Carrell et al., 2003)
3. Edad del varón >40 años (Singh et al., 2003)
4. Varicocele (Smith et al., 2006)
5. Episodio de fiebre alta en los últimos tres meses (Evenson et al., 2000)
6. Cuando se va a congelar semen antes de hacer una vasectomía
7. Cuando se va a congelar semen previo a tratamientos de quimio/radioterapia
2. ¿Si una muestra de semen de un paciente tiene un valor de fragmentación elevado, es ésta una condición permanente o pudiera ser que otras muestras tuviesen valores más bajos?
Si bien en donantes fértiles los valores de fragmentación suelen ser relativamente bajos y la variación en el grado de fragmentación de DNA en muestras de semen obtenidas en diferentes ciclos espermatogénicos es también baja (Evenson et al., 1991), en pacientes de infertilidad, el grado de fragmentación puede mostrar fluctuaciones significativas (Álvarez et al., 2004). Estas fluctuaciones pudieran estar relacionadas con alteraciones en el control de la espermiogénesis (p.e., varicocele, proceso inflamatorio subclínico, proceso febril) o con factores desencadenantes físico-químicos que induzcan apoptosis o activen endonucleasas espermáticas (p.e., proceso febril, tóxicos medioambientales) que podrían inducir fragmentación del DNA en los espermatozoides después de la espermiación (Evenson et al., 2000). Una vez que haya cesado el estímulo desencadenante, el grado de fragmentación de DNA se normalizaría en el ciclo espermatogénico siguiente. Por ello, en aquellos casos en los que la muestra de semen analizada tenga un valor por encima del umbral de normalidad del test utilizado, deberían recogerse varias muestras de semen, analizar el grado de fragmentación de DNA, criopreservarlas, y utilizar únicamente aquellas muestras con valores de fragmentación dentro de la normalidad. Esto sería particularmente útil en aquellos casos en los que las muestras de semen se vayan a congelar, p.e., pacientes que van a ser sometidos a quimio o radioterapia.
3. ¿Qué factores pueden causar fragmentación de DNA?
Una gran variedad de factores han sido asociados con la inducción de daño de DNA en los espermatozoides. Estos factores incluyen procesos inflamatorios agudos y crónicos, el estrés oxidativo, varicocele, fiebre alta, exposición del testículo a temperaturas >36ºC (Banks et al., 2005), tóxicos medioambientales, quimioterapia, radiación ionizante, edad >40 años, consumo excesivo de cafeína (= o > tres tazas de café al día) (Schmid et al., Hum Reprod 2006, in press).
4. ¿Si el valor del test es normal, garantiza esto un alto nivel de fertilidad?
No necesariamente. La calidad del DNA pudiera ser óptima y sin embargo otros parámetros espermáticos u ovocitarios podrían estar también afectados. Como se ha indicado anteriormente, los tests de fragmentación pueden tener un alto valor predictivo negativo pero no positivo. Las tasas de fecundación e implantación, además de depender de la calidad genómica del espermatozoide que haya fecundado al ovocito, van a depender también de otros parámetros como son la calidad citoplasmática y genómica del ovocito y la receptividad del endometrio.
5. ¿Qué se hace cuando los valores de fragmentación de DNA obtenidos en varias muestras de semen son consistentemente altos? ¿tendría que recurrirse a semen de donante?
No. Muestras de semen con un valor de fragmentación por encima del umbral de normalidad pueden producir embarazos a término, independientemente de la edad de la mujer. Esto es particularmente cierto cuando se utilizan tests con un valor predictivo bajo. Estudios recientes muestran que muestras de semen con valores de fragmentación relativamente altos medidos por el test SCSA (Gandini et
al., 2004; Bungum et al., 2004) o por el test SCD (Muriel et al., 2006) pueden resultar en embarazos viables a término. De ahí que se recomiende el uso de tests de fragmentación que miden daño real, como el test TUNEL, en combinación con tests que miden daño secundario de DNA como la presencia de 8-OH-2´-deoxiguanosina.
6. ¿Por qué muestras de semen con valores de fragmentación relativamente altos pueden dar lugar a un embarazo a término?
El valor predictivo de un test de fragmentación de DNA va a depender de varios factores. Estos factores incluyen:
1. Fragmentación de DNA de cadena sencilla vs. cadena doble. En general el daño de cadena sencilla es más fácil de reparar por el ovocito que el de cadena doble.
2. Porcentaje de espermatozoides que están afectados. Cuanto más alto sea el porcentaje de espermatozoides con DNA fragmentado menor la probabilidad de que un espermatozoide con DNA intacto pueda fecundar al ovocito.
3. Grado de fragmentación de DNA por espermatozoide. Cuanto mayor sea el grado de fragmentación del DNA nuclear del espermatozoide, menor la probabilidad de que el ovocito lo pueda reparar.
4. Si existe únicamente daño primario o si existe daño combinado primario y secundario. Por ejemplo, en casos de apoptosis durante la espermatogénesis o roturas de DNA producidas este tipo de daño no suele estar asociado a daño secundario. Por el contrario, en casos de daño post-testicular inducido por el radical hidroxilo o en casos de exposición a radiación ionizante, el daño primario (que suele
ser de cadena doble) casi siempre está asociado a daño secundario.
5. Si el daño afecta intrones o exones. Más del 90% del DNA está formado por intrones. Por lo tanto, la probabilidad de que el daño de DNA afecte a exones, que son las secuencias del DNA que codifican proteínas, es relativamente baja.
6. Capacidad del ovocito de reparar daño de DNA en el espermatozoide que lo ha fecundado. Esta capacidad puede variar de un ovocito a otro en cohortes de ovocitos obtenidos en ciclos de in vitro. Desdichadamente, la capacidad de reparación del ovocito no se puede determinar in vitro, ya que esto afectaría su viabilidad.
7. Número de ovocitos. En ciclos de in vitro en parejas donde el nivel de fragmentación de DNA en semen, medido por TUNEL, es relativamente alto, la probabilidad de que se produzcan embriones viables y un embarazo a término va a depender, al menos en parte, del número de ovocitos obtenidos y de su capacidad de reparar daño de DNA en el espermatozoide. Si se dispone de seis ovocitos en metafase II (Fig. 1), la probabilidad de que se produzca un embarazo va a ser mayor que si se dispone de tan sólo tres ovocitos (Fig. 2). En este último caso, la probabilidad de que un espermatozoide con el DNA intacto fecunde un óvulo normal o de que un espermatozoide con DNA fragmentado fecunde un óvulo con una alta capacidad reparadora va a ser mucho menor que en el primer caso. Si bien en el primer caso, como se indica en la Fig. 1, podrían producirse dos embriones grado I, que podrían dar lugar a un embarazo, ello no quiere decir de que la fragmentación de DNA no esté asociada a un mal pronóstico en TRA, ya que de haber menos ovocitos como en el caso ilustrado en la Fig. 2, podría no producirse ningún embrión viable y, por lo tanto, no se produciría un embarazo. Por lo tanto, el número de ovocitos disponibles es un factor importante que va a influir en el valor predictivo de tests de fragmentación de DNA. Esto mismo se aplicaría a ciclos de coito dirigido e IAC, donde el número de ovocitos que se obtienen habitualmente es de uno y dos, respectivamente. Aquí las tasas de embarazo serían más bajas cuando el grado de fragmentación es relativamente alto, ya que habitualmente se desarrollan de uno a dos folículos. Por lo tanto, uno esperaría que el valor predictivo negativo de tests de fragmentación, especialmente aquellos que miden daño real como el test TUNEL, sea más alto en ciclos de IAC. Esto ha sido confirmado en un estudio donde se encontró una correlación significativa entre el grado de fragmentación de DNA en semen medido por el test TUNEL y las tasas de embarazo en ciclos de IAC (Duran et al., 2002).
Para concluir, podríamos decir que el valor predictivo de un test de fragmentación, IItest, es el sumatorio de varios factores: IItest = II(1) + II(2) + II(3)…….+II(n). Algunos de estos factores pueden medirse y otros no como, por ejemplo, si el daño afecta a intrones o exones o la capacidad reparadora del ovocito.

7. ¿Qué test de fragmentación de DNA es el más recomendable?
Se recomienda utilizar tests que midan daño primario y daño secundario. Para determinar daño primario se recomienda el uso de tests de fragmentación que miden daño real como el test TUNEL, ya sea por microscopía o por citometría de flujo. Los kits comerciales que existen para el test TUNEL son sencillos, poco costosos, permiten el uso de microscopía de campo claro o de fluorescencia, dependiendo de si la sonda va conjugada a peroxidasa o fluoresceína, respectivamente (Roche Diagnostics, Mannheim, Germany) y, por lo tanto, pueden realizarse en cualquier laboratorio. No se recomienda el uso de tests como el SCSA o el test SCD, incluyendo el Halosperm kit, ya que estos tests miden daño potencial o susceptibilidad a la desnaturalización del DNA y tienen un valor predictivo relativamente bajo en TRA. Para medir daño secundario se recomienda la determinación de 8-OH-2´-deoxiguanosina utilizando el kit de ELISA de Oxford Biomedical (Oxford Biomedical Research, USA).
Referencias
Aitken RJ, Gordon E, Harkiss D, Twigg JP, Milne P, Jennings Z, Irvine DS. Relative impact of oxidative stress on the functional competence and genomic integrity of human spermatozoa. Biol Reprod 1998; 59:1037-46.
Álvarez JG, Ollero M, Larson-Cook KL, Evenson DP. Selecting cryopreserved semen for assisted reproductive techniques based on the level of sperm nuclear DNA fragmentation resulted in pregnancy. Fertil Steril 2004; 81:712-3.
Banks S, King SA, Irvine DS, Saunders PT. Impact of mild scrotal heat on DNA integrity in murine spermatozoa. Reproduction 2005; 129:505-14.
Benchaib M, Braun V, Lornage J, Hadj S, Salle B, Lejeune H, Guerin JF. Sperm DNA fragmentation decreases the pregnancy rate in an assisted reproductive technique. Hum Reprod 2003; 18:1023-8.
Billig H, Chun SY, Eisenhauer K, Hsueh AJ. Gonadal cell apoptosis hormoneregulated cell demise. Hum. Reprod. Update 1996; 2:103-117.
Brandiff B and Pedersen RA. Repair of the ultraviolet-irradiated male genome in fertilized mouse eggs. Science 1981; 211:1431-1433.
Britan A, Maffre V, Tone S, Drevet JR. Quantitative and spatial differences in the expression of tryptophan-metabolizing enzymes in mouse epididymis. Cell Tissue Res 2006; 2:1-10.
Bungum M, Humaidan P, Spano M, Jepson K, Bungum L, Giwercman A. The predictive value of sperm chromatin structure assay (SCSA) parameters for the outcome of intrauterine insemination,
IVF and ICSI. Hum Reprod 2004; 19:1401-8.
Burrello N, Arcidiacono G, Vicari E, Asero P, Di Benedetto D, De Palma A, Romeo R, D´Agata R, Calogero AE. Morphologically normal spermatozoa of patients with secretory oligo-astheno- teratozoospermia have an increased aneuploidy rate. Hum Reprod 2004; 19:2298-302.
Carrell DT, Liu L, Peterson CM, JOnes KP, Hatasaka HH, Erickson L, Campbell B. Sperm DNA fragmentation is increased in couples with unexplained recurrent pregnancy loss. Arch Androl 2003; 49:49-55.
Duran EH, Morshedi M, Taylor S, Oehninger S. Sperm DNA quality predicts intrauterine insemination outcome: a prospective cohort study. Hum Reprod 2003; 17:3122-8.
Evenson DP, Darzynkiewicz Z, and Melamed, MR. Relation of mammalian sperm heterogenity to fertility. Science 1980; 210:1131-133.
Evenson DP and Melamed MR. Rapid analysis of normal and abnormal cell types in human semen and testis biopsies by flow cytometry. J. Histochem Cytochem 1983; 31:248- 53.
Evenson DP, Higgins PJ, Grueneberg D, Ballachey BE. Flow cytometric analysis of mouse spermatogenic function following exposure to ethylnitrosourea. Cytometry 1985; 6:238-53.
Evenson DP, Jost LK, Baer RK, Turner TW, Schrader SM. Individuality of DNA denaturation patterns in human sperm as measured by the sperm chromatin structure assay. Reprod. Toxicol. 1991; 5:115-25.
Evenson DP and Jost, LK. Sperm chromatin structure assay: DNA denaturability. In: Methods in cell biology, Vol 42., Flow Cytometry (Darzynkiewicz, Z., Robinson, J.P., and Crissman, H.A. (eds) Academic Press, Orlando, 1994; pp:159
Evenson DP. Loss of livestock breeding efficiency due to uncompensable sperm nuclear defects. Reprod Fertil Dev 1999a; 11:1-15.
Evenson DP, Jost LK, Marshall D, Zinaman MJ, Clegg E, Purvis K, de Angelis P, Claussen OP. Utility of the sperm chromatin structure assay as a diagnostic and prognostic tool in the human fertility clinic. Hum Reprod 1999b; 14:1039-49.
Evenson DP, Jost LK, Corzett M, Balhorn R. Characteristics of human sperm chromatin structure following an episode of influenza and high fever: a case study. J Androl 2000; 21:739-46.
Fernández JL., Goyanes VJ., Ramiro- Díaz J., Gosalves J. Application of FISH for in situ detection and quantification of DNA breakage. Cytogenet Cell Genet 1998; 82:251-56.
Fernández JL, Vázquez-Gundin F, Delgado A, Goyanes VJ, Ramiro-Díaz J, de la Torre J, Gosalvez J. DNA breakage detection-FISH (DBD-FISH) in human spermatozoa: technical variants evidence different structural features. Mutat Res 2000; 453:77- 82.
Fernández JL, Muriel L, Rivero MT, Goyanes V, Vázquez R, Álvarez JG. The sperm chromatin dispersion test: a simple method for the determination of sperm DNA fragmentation. J Androl 2003; 59-66.
Gandini L, Lombardo F, Paoli D, Caruso F, Eleuteri P, Leter G, Ciriminna R, Culasso F, Dondero F, Lenzi A, Spano M. Full-term pregnancies achieved with ICSI despite high levels of sperm chromatin damage. Hum Reprod 2004; 19:1409-17.
Genesca A, Caballin MR, Miro R, Benet J, Germa JR, Egozcue J. Repair of human sperm chromosome aberrations in the hamster egg. Hum. Genet 1992; 82:181-86.
Gorczyca W, Gong J, Darzynkiewicz Z. Detection of DNA strand breaks in individual apoptotic cells by the in situ terminal deoxynucleotidyl transferase and nick translation assays. Cancer Res 1993; 53:945-951.
Greco E, Scarselli F, Iacobelli M, Rienzi L, Ubaldi F, Ferrero S, Franco G, Anniballo N, Mendoza C, and Tesarik J. Efficient treatment of infertility due to sperm DNA damage by ICSI with testicular spermatozoa. Hum. Reprod. 2005; 20:226-30.
Greco E, Iacobelli M, Rienzi L, Ubaldi F, Ferrero S, Tesarik J. Reduction of the incidence of sperm DNA fragmentation by oral antioxidant treatment. J Androl 2005; 26:349-53.
Hughes CM, Lewis SE, McKelvey-Martin VJ, Thompson W. A comparison of baseline and induced DNA damage in human spermatozoa from fertile and infertile men using a modified comet assay. Mol Hum Reprod 1996; 2:613-19.
Larson K., DeJonge C., Barnes A. Jost L, and Evenson DP. Relationship between assisted reproductive techniques (ART) outcome and status of chromatin integrity as measured by the Sperm Chromatin Structure Assay (SCSA). Hum Reprod 2000; 15:1717-22.
Larson-Cook KL, Brannian JD, Hansen KA, Kasperson KM, Aamold ET, Evenson DP. Relationship between the outcomes of assisted reproductive techniques and sperm DNA fragmentation as measured by the sperm chromatin structure assay. Fertil Steril 2003; 80:895-902.
Lopes, S., Jurisicova, A., Sun, J.G. et al. Reactive oxygen species: potential cause for DNA fragmentation in human spermatozoa. Hum. Reprod. 1998; 13:896-900.
Lopes S, Sun JG, Jurisicova A, Meriano J, Casper RF. Sperm deoxyribonucleic acid fragmentation is increased in poor-quality semen samples and correlates with failed fertilization in intracytoplasmic sperm injection. Fertil Steril 1998; 69:528-32.
Manicardi GC, Bianchi PG, Pantano S, Azzoni P, Bizzaro D, Bianchi U, Sakkas D. Presence of endogenous nicks in DNA of ejaculated human spermatozoa and its relationship to chromomycin A3 accessibility. Biol Reprod 1995; 52:864-67.
McPherson SMG and Longo FJ. Localization of DNAse I-hypersensitive regions during rat spermatogenesis: stage-dependent patterns and unique sensitivity of elongating spermatids. Mol Reprod. Dev 1992; 31:268-79.
McPherson SMG and Longo FJ. Nicking of rat spermatid and spermatozoa DNA: possible involvement of DNA tropoisomerase II. Devel Biol 1993a; 158:122-30.
McPherson SMG and Longo FJ. Chromatin structure-function alterations during mammalian spermatogenesis: DNA nicking and repair in elongating spermatids. Eur J Histochem 1993b; 37:109-128.
Matsuda Y. and Tobari I. Chromosomal analysis in mouse eggs fertilized in vitro with sperm exposed to ultraviolet light and methyl and ethyl methansulfonate. Mutation Res 1988; 198:131- 44.
Mortimer D, Menkveld R. Sperm morphology assessment—historical perspectives and current opinions. J Androl 2001; 22:192-205.
Muriel L, Meseguer M, Fernandez JL, Alvarez JG, Remohi J, Pellicer A, GArrido N. Value of the sperm chromatin dispersion test in predicting pregnancy outcome in intrauterine insemination: a blind prospective study. Hum Reprod 2006; 21:738- 44.
Obasaju M, Kadam A, Sultan K, Fateh M, Munne S. Sperm quality may adversely affect the chromosome constitution of embryos that result from intracytoplasmic sperm injection. Fertil Steril 1999; 72:1113-25.
Ollero M, Gil-Guzmán E, López MC, Sharma RK, Agarwal A, Larson K, Evenson D, Thomas AJ Jr, Álvarez JG. Characterization of subsets of human spermatozoa at different stages of maturation: implications in the diagnosis and treatment of male infertility. Hum Reprod. 2001; 16:1912-21.
Parinaud J, Mieusset R, Vieitez G, Labal B, Richoilley G. Influence of sperm parameters on embryo quality. Fertil Steril 1993; 60:888-892.
Pentikainen V, Erkkila K, and Dunkel L. Fas regulates germ cell apoptosis in the human testis in vitro. Am. J. Physiol. 1999; 276:E310-E316.
Sailer BL, Sarkar LJ, Bjordahl JA, Jost LK, Evenson DP. Effects of heat stress on mouse testicular cells and sperm chromatin structure. J Androl 1997; 18:294-301.
Sakkas D, Mariethoz E, Manicardi G, Bizzaro D, Bianchi PG, Bianchi U. Origin of DNA damage in ejaculated human spermatozoa. Revi Reprod 1999; 4:31-37.
Sakkas D, Moffatt O, Manicardi GC, Mariethoz E, Tarozzi N, Bizzaro D. Nature of DNA damage in ejaculated human spermatozoa and the possible involvement of apoptosis. Biol Reprod. 2002; 66:1061-7.
Schmid T, Eskenazi B, Baumgartner A, Marchetti F, Young S, Weldon R, Anderson D, Wyrobek A. The effects of male age on sperm DNA damage in healthy non-smokers. Hum Reprod, [in press].
Sergerie M, Laforest G, Bujan L, Bissonnette F, Belau G. Sperm DNA fragmentation: threshold value in male fertility. Hum Reprod 2005; 20:3446-51.
Singh NP, Muller CH, Berger RE. Effects of age on double-strand breaks and apoptosis in human sperm. Fertil Steril 2003; 80:1420-30.
Smith R, Kaune H, Parodi D, Madariaga M, Rios R, Morales I, Castro A. Increased sperm DNA damage in patients with varicocele: relationship with seminal oxidative stress. Hum Reprod 2006; 21:986-93.
Steele EK, McClure N, Maxwell RJ, Lewis SEM. A comparison of DNA damage in testicular and proximal epididymal spermatozoa in obstructive azoospermia. Mol Hum Reprod 1999; 9:831-5.
Suganuma R, Yanagimachi R, Meistricht M. Decline in fertility of maouse sperm with abnormal chromatine during epididymal passage as revealed by ICSI. Hum Reprod 2005; 20:3101-08.
Tesarik J, Greco E, Mendoza C. Late, but not early, paternal effect on human embryo development is related to sperm DNA fragmentation. Hum Reprod 2004; 19:611-5.
Tomlinson MJ, Moffatt O, Manicardi GC, Bizzaro D, Afnan M, Sakkas D. Interrelationships between seminal parameters and sperm nuclear DNA damage before and after density gradient centrifugation: implications for assisted conception. Hum Reprod 2001; 16:2160-5.
Twigg, JP, Irvine, DS and Aitken, RJ. Oxidative damage to DNA in human spermatozoa does not preclude pronucleus formation at intracytoplasmic sperm injection. Hum Reprod 1998; 13:1864-71.