PAPEL DE LA INTERACCIÓN CELULAR EN FOLICULOGÉNESIS
Paloma Sánchez-Aparicio1, Ricardo Ramos2 y Jorge Cuadros1
1Clínica de Medicina de la Reproducción y Ginecología, Fiv Madrid, Madrid. 2Unidad de Genómica, Parque Científico, Universidad Autónoma, Madrid.
Resumen: El proceso de crecimiento y maduración folicular que conlleva a la formación de gametos femeninos funcionales se denomina foliculogénesis. Este proceso está estrictamente regulado de forma endocrina y paracrina. Además, las interacciones celulares, tanto célula-célula como célula-MEC (matriz extracelular), desempeñan un papel fundamental. En concreto, el correcto diálogo intercelular controla la adecuada maduración oocitaria. Este artículo es una revisión de los principales mecanismos de interacción celular así como las familias de moléculas de adhesión implicadas.
Palabras clave: adhesión, interacción celular, foliculogénesis
The role played by cell interaction in folliculogenesis
Summary: The process of follicular growing and maturation which leads to the formation of functional female gametes is named folliculogenesis and is subject to a strict endocrine and paracrine regulation. In addition, cell interactions (cell-cell and cell-ECM) play an essential role in this process. In particular, the proper maturation of oocytes is controlled by a correct intercellular dialogue. This review focuses on the main mechanisms of cell interaction as well as the most important families of adhesion molecules involved.
Key words: adhesion, cell-cell interaction, folliculogenesis
Regulación funcional del ovario
El proceso de foliculogénesis tiene lugar en los ovarios y asegura la formación de gametos femeninos funcionales (oocitos) que están diseñados para transmitir la carga genética a la siguiente generación. Los gametos, que son haploides, darán lugar a un zigoto diploide cuyas células proliferarán y terminarán por diferenciarse.
En la cavidad peritoneal se encuentran los ovarios cuya arquitectura tisular es sencilla: en cada uno de ellos se distingue una región cortical y un estroma medular central. Asimismo, el tejido ovárico está constituido por células germinales, células epiteliales superficiales circundantes y células de estroma -células de tejido conectivo y productoras de hormonas-.Las células germinales darán lugar a los oocitos mientras que el resto de las células están implicadas en la creación del microambiente adecuado que garantiza el correcto desarrollo oocitario. Desde la pubertad hasta la menopausia, el ovario se caracteriza por tener una estructura dinámica donde los folículos ováricos están en continuo desarrollo. Los folículos se sitúan en la región cortical del ovario y están en evolución continua a partir del estadio primordial. Los folículos primordiales se forman, en su totalidad, durante el desarrollo prenatal y están constituidos por un oocito detenido en profase de la primera división meiótica y una capa de células somáticas planas circundantes (Lei et al., 2006). Durante un periodo variable, los folículos primordiales están en estado de quiescencia (Fortune et al., 2000). La mayoría permanecen en este estado hasta que degeneran o se activan por señales específicas que les hacen entrar en fase de crecimiento, sufriendo entonces un dramático proceso de proliferación y diferenciación (Figura 1).
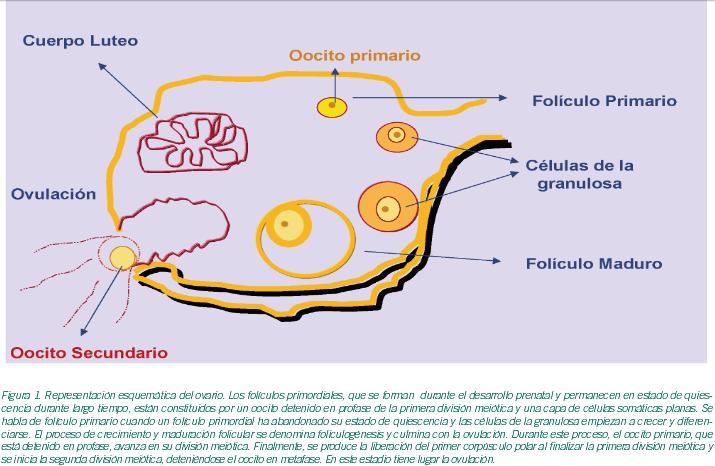
Aunque los mecanismos concretos que bloquean o inician el crecimiento de los folículos primordiales en estado quiescente no se han establecido con exactitud, el conocimiento de los genes que se expresan podría ayudar a entender los mecanismos de regulación de este proceso (Kocabas et al., 2006; Yoon et al., 2006).
Dicho proceso de crecimiento y maduración folicular constituye la foliculogénesis y culmina con la ovulación en cada ciclo ovárico durante el periodo reproductivo. El ovario además de estar implicado en la producción de gametos desempeña una función endocrina esencial. La producción de hormonas esteroideas está, a su vez, implicada en la propia generación de gametos por parte del ovario. Dicha producción hormonal está bajo el control del eje hipotálamo–hipofisario, con mecanismos de control estrictamente regulados (regulación endocrina).
Los ovarios disponen además de sistemas de regulación locales; esto es, células cuyas secreciones se autorregulan (regulación autocrina); y células cuyas secreciones se producen para intervenir en el control de células vecinas de la misma gónada (regulación paracrina). Por tanto, las células de los ovarios responden al ambiente externo y a las señales de otras células; la correcta producción de gametos depende de la acción combinada de este complejo sistema de secreciones. Las señales que reciben las células del tejido ovárico son trasladadas a diferentes partes de la propia célula, incluido el núcleo (Pangas et al., 2000).
Estas vías de transducción de señales acaban regulando el crecimiento, la supervivencia, la diferenciación, la migración y el metabolismo celular, y por tanto la transcripción de un número considerable de genes. En todo este proceso, los oocitos avanzan en su estado madurativo y simultáneamente las células de la granulosa desarrollan capacidades esteroidogénicas específicas.
En etapas iniciales, se han descrito numerosos factores intra-ovario, miembros de distintas familias de factores de crecimiento que influyen en el crecimiento y la maduración folicular a través de la señalización paracrina (Hernández et al., 1991, 1992; Matzuk et al., 2002). Sin embrago, los estadios tardíos del desarrollo folicular son dependientes de las hormonas hipofisarias FSH (hormona folículo estimulante) y LH (hormona luteinizante) (Shimada et al., 2002); la primera es el mayor promotor de la maduración folicular aumentando la proliferación de las células de la granulosa y regulando la producción de la hormona estradiol (hormona esteroidea), la segunda juega un papel destacado en estadios avanzados del desarrollo folicular, estimulando la maduración del oocito, la ovulación y el proceso de luteinización. A su vez, los factores de crecimiento producidos por los propios folículos son capaces de modular, amplificando o atenuando, la acción de la hormona FSH (Adashi et al., 1993).
En definitiva, el proceso de la foliculogénesis está regulado por mecanismos intra-ovario y endocrinos que coordinan el proceso de crecimiento, proliferación y diferenciación celular (Peramo et al., 2002). En el interior del folículo las interacciones paracrinas entre el oocito y las células de la granulosa circundantes son críticas para una correcta función celular y un adecuado desarrollo oocitario (Guigon et al., 2006).
De especial relevancia en el crecimiento del oocito son los procesos mediados por la interacción Kit-kit ligand (Driancourt et al., 2000), así como Kit ligand a través de la interacción con GDF9 y BMP15 (Hutt et al., 2006; Thomas et al., 2006; Miyoshi et al., 2006).
Mecanismos de interacción celular
Los procesos de interacción celular aseguran el mantenimiento de la arquitectura tisular y garantizan el correcto desarrollo funcional de los órganos. Dichos procesos incluyen las interacciones célula-célula y célula-MEC (matriz extracelular) y pueden tener lugar vía estructuras de gran complejidad (cell junction) o a través de simples contactos celulares (non cell junction).Las cell junctions incluyen especializaciones de la membrana celular del tipo de las tight junctions, adherens junctions, gap junctions, desmosomas, hemidesmosomas o contactos focales; mientras que las segundas son meros contactos célula- célula y célula-MEC de gran sencillez estructural y dinamismo (Figura 2).
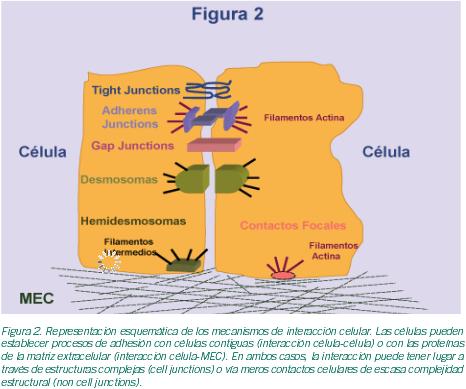
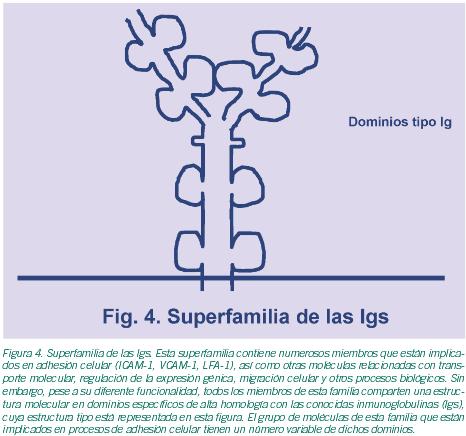
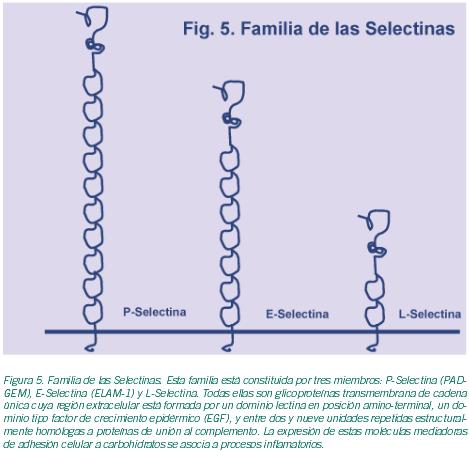
En los ovarios, y en particular en los folículos, se dan la mayor parte de los mecanismos de interacción celular mencionados. De todos ellos, las gap junctions (GJ) han sido descritas como elementos clave en el proceso de la foliculogénesis. Las GJ son agrupaciones de canales intercelulares que permiten una comunicación citoplasmática directa entre células adyacentes (Evans et al., 2002). Estos canales están, a su vez, formados por subunidades de proteínas denominadas conexinas (Cxs); y permiten el paso de pequeñas moléculas -iones, metabolitos, segundos mensajeros- haciendo posible de este modo la sincronización del comportamiento celular. Estas estructuras permiten la coordinación entre las propias células de la granulosa, y de éstas con el oocito; de ahí la relevancia de las GJ en la maduración oocitaria (Kidder et al., 2002; Wright et al., 2001). Está ampliamente admitido que las interacciones celulares que se producen en el propio folículo ovárico desempeñan un papel esencial para la correcta maduración oocitaria. Así pues, el complejo diálogo intercelular resulta crucial para la regulación de este proceso. Además de las ya citadas Cxs, existen otras moléculas implicadas en estos procesos. Estas últimas pertenecen a distintas familias de moléculas de adhesión que incluyen la superfamilia de las Integrinas, la superfamilia de las Ig(s), la familia de las Selectinas, la familia de las Cadherinas, la familia de los Proteoglicanos y las Sialomucinas (Tabla I; Figuras 3-6); y están formando parte de las complejas estructuras mencionadas o de simples contactos celulares, según los casos.Procesos de interacción célula-célula implicados en foliculogénesis. Durante el proceso de foliculogénesis, las interacciones célula-célula mediadas a través de las GJ, en particular las GJ de conexina 43 (Cx43), desempeñan un papel fundamental (Tabla II).


La Cx43 es un miembro de la familia de las Cxs que se expresa mayoritariamente en el folículo ovárico y cuyo papel es clave en foliculogénesis. El estudio llevado a cabo con ratones knockout para Cx43 pone de manifiesto que la presencia de esta molécula resulta imprescindible para mantener la comunicación entre las células de la granulosa que rodean al oocito y sostener, por tanto, la proliferación de estas células y la maduración folicular; en ausencia de Cx43 los folículos se detienen en estadio preantral temprano y los oocitos resultan incompetentes (Juneja et al., 1999; Ackert et al., 2001).
Las GJ de Cx43 existen ya en folículos primordiales, la expresión de Cx43 se acumula coincidiendo con el crecimiento folicular, y dicha expresión está sujeta a modulación durante el ciclo ovárico a través de las hormonas FSH y LH: la primera estimula la síntesis de Cx43 y la amplificación de canales intercelulares funcionales, mientras que la segunda, en una respuesta inmediata, interrumpe la comunicación célula-célula por la fosforilación y cambio conformacional de la molécula de Cx43 que conduce al cierre de los canales de las GJ (Granot et al., 2002, Yogo et al., 2002; Yogo et al., 2006).

La hormona LH tiene, por tanto, un efecto inmediato que es seguido por una respuesta tardía que conduce a la eliminación de la proteína por estimulación de la reducción de los niveles de mRNA; y a la desaparición final de las GJ de Cx43 (Gittens et al., 2005). Algunos autores han propuesto además que el efecto de las hormonas FSH y LH está mediado por las hormonas esteroideas sintetizadas en el propio ovario, aunque se desconocen con exactitud los mecanismos implicados en este proceso.
Además, se ha descrito la expresión de otros miembros de la familia de las Cx(s), entre ellos, Cx26, Cx30, Cx32, Cx37, Cx40 y Cx45 en tejido ovárico de diferentes especies de mamíferos (Goodenough et al., 1999; Teilmann et al., 2005; Simon et al., 2006). Aunque las células de la granulosa se comunican entre ellas a través de GJ de Cx43, éstas se comunican con el oocito a través de GJ de Cx37 (Gittens et al., 2005).
La Cx37 desempeña un papel esencial para el desarrollo folicular, la maduración del oocito y la ovulación; en ausencia de Cx37, los folículos no llegan a folículo antral y es causa de infertilidad en ratones (Simon et al., 2002) (Tabla II). La comunicación celular a través de las GJ de Cx37 garantiza el paso de nutrientes y segundos mensajeros al interior del oocito desde las células de la granulosa y viceversa. Está ampliamente admitido que la comunicación oocito-células de la granulosa es bidireccional, de manera que el oocito recibe información del exterior pero a su vez la información procedente del oocito condiciona la evolución de las células de la granulosa que lo rodean (Guigon et al., 2006).
Existen otras moléculas implicadas en contactos célula-célula que desempeñan un papel relevante en la arquitectura y el desarrollo folicular (Tabla II). Este es el caso de las moléculas de adhesión dependientes de calcio, las cadherinas, que regulan la formación de las cell junctions, así como el establecimiento de la polaridad celular (Schuldt et al., 2005).
La implicación de las cadherinas en la remodelación tisular sugiere una implicación inequívoca en la reorganización celular del ovario en su desarrollo postnatal. Así pues, cambios de expresión de las cadherinas N y E (Cad-N, Cad-E) se han descrito coincidiendo con el crecimiento ovárico y la foliculogénesis (Machell et al., 2000), así como cambios en el perfil de distribución de otras cadherinas (Machell et al., 2003; Ziv et al., 2002).
La Cad-E está presente entre las células de la granulosa y forma parte de las estructuras tipo cell junctions que garantizan un estrecho contacto celular; y desempeña un papel importante en el estadio preantral tardío y antral (Machell et al., 2003). Además, el análisis de los ratones knockout para Cad-E demuestra el papel esencial que desempeña esta molécula en el desarrollo embrionario temprano puesto que ninguno de los embriones sin Cad- E llega a alcanzar el estadio de blastocisto (Larue et al., 1996).
Finalmente, otras moléculas implicadas en interacciones célula-célula, como por ejemplo ICAM-1, VCAM-1 y L-Selectina (Campbell et al., 1995) o incluso LFA-1 (Lu et al., 2002) también se han detectado en oocitos aunque su función exacta se desconoce hasta el momento (Tabla II).
Procesos de interacción célula-MEC implicados en foliculogénesis
Los procesos de interacción celular engloban, además de las interacciones célula-célula, las interacciones que las células establecen con proteínas de la matriz extracelular (MEC). La MEC está formada por un conjunto de macromoléculas como laminina, fibronectina, perlecan, nidogen y distintos tipos de colágeno, que participan en procesos biológicos como migración celular, proliferación, crecimiento y desarrollo. En el ovario, la MEC constituye la lámina folicular basal y está presente entre las células foliculares y en el fluido folicular, y juega un papel relevante en la función ovárica según demuestran los numerosos trabajos publicados en los últimos años (Clavero et al., 2004; Ahmed et al., 2005; Irving-Rodgers et al., 2005, 2006; Shozu et al., 2005; Russel et al., 2006; Curry et al., 2006; Berkholtz et al., 2006a,b).
En el ovario, los folículos contienen diferentes tipos celulares y compartimentos separados que cambian a lo largo del proceso de crecimiento y desarrollo folicular.
Aunque la cantidad relativa y la distribución de las proteínas de matriz dentro del folículo en desarrollo no se sabe con exactitud (Berkholtz et al., 2006a) parece claro que son capaces de promover o inhibir procesos celulares como la proliferación, la diferenciación y la supervivencia celular que ocurren durante el desarrollo folicular.
Los componentes de la matriz extracelular varían en los distintos compartimentos foliculares, teniendo dichos componentes una implicación diferencial en los procesos de desarrollo folicular y atresia celular (Irving-Rodgers et al., 2006). Además, la composición de la matriz no sólo varía por los cambios en la síntesis de nuevos componentes, sino por la degradación proteolítica de las proteínas debida a la acción de enzimas como las metalloproteasas, activadores del plasminógeno y proteínas ADAMTS (Shozu et al., 2005; Curry et al., 2006).
En la actualidad se ha demostrado de forma inequívoca el papel de la MEC en la regulación de los procesos celulares que tienen lugar en el ovario (Berkholtz et al., 2006b), así como la activación transcripcional de algunos genes de MEC en la transición de folículos primordiales a folículos primarios, o incluso la inhibición de un número elevado de otros genes de MEC en el mismo estadio de desarrollo evolutivo (Yoon et al., 2006). Por tanto, la formación y el desarrollo, así como la ovulación y la regresión de los folículos se asocian a una considerable remodelación tisular, tal que los distintos constituyentes de la MEC -a través de sus receptores específicos de membrana- estarían directamente implicados en tales procesos (Ahmed et al., 2005; Irving-Rodgers et al., 2005, 2006; Shozu et al., 2005; Russel et al. 2006).
Las moléculas que se expresan en la superficie celular y actúan como receptores de las proteínas de MEC pertenecen, a su vez, a distintas familias de moléculas de adhesión (ver Tabla I). La familia más numerosa y más ampliamente descrita es la superfamilia de las integrinas (Figura 3), cuyos miembros pueden formar parte de estructuras complejas o participar en meros contactos celulares.
Las integrinas facilitan la adhesión de las células a la MEC y activan además los mecanismos de señalización que regulan la reestructuración del citoesqueleto, el comportamiento celular y la síntesis de proteínas. Diversos trabajos han demostrado la presencia de distintas subunidades a y b de las integrinas en el ovario (Burns et al., 2002), en oocitos (Pate et al., 2006) y en células de la granulosa (Clavero et al., 2004).
En concreto, la subunidad a6 de las integrinas se ha descrito en células de la granulosa de folículos preovulatorios (Nakamura et al., 1997) y desempeña un papel clave en foliculogénesis (Honda et al., 1995; Fujiwara et al., 1996) (Tabla III). Asimismo, la integrina a6b1 asociada a CD9 está implicada en la regulación de la posterior luteinización de estas células a través de su ligando de MEC, la laminina (Takao et al., 1999).

A su vez, miembros de otras familias de moléculas de adhesión como CD44 (HCAM), receptor del ácido hialurónico y miembro de la familia de los proteoglicanos, está implicado tanto en procesos de interacción célula-célula como célula-MEC, y también se expresa en oocitos (Lu et al., 2002) así como en complejos cúmulo-oocito (Assou et al., 2006; Sato et al., 2005)
(Tabla III).
En conclusión, la foliculogénesis es un proceso de una extraordinaria complejidad, que por tanto está sujeto a una estricta regulación y en el que las interacciones celulares desempeñan un papel fundamental. Numerosos estudios desarrollados en animales experimentales demuestran que el estrecho diálogo intercelular garantiza la correcta consecución de la maduración oocitaria.
En este artículo se hace una revisión de las numerosas moléculas implicadas tanto en contactos célula-célula como célula-MEC, así como las diferentes estructuras de las que forman parte, que gobiernan por mecanismos diversos el proceso de la foliculogénesis.
En conclusión, una sincronizada interacción bidireccional entre las células somáticas y las células germinales asegura el proceso normal de la foliculogénesis y su adecuada duración.
Financiación: Este trabajo ha sido financiado en su totalidad de forma privada por la gerencia de la Clínica de Medicina de la Reproducción y Ginecología, FivMadrid.
Referencias
Ackert CL, Gittens JE, O´Brien MJ, Eppig JJ, Kidder GM. Intercellular communication via connexin 43 gap junctions is required for ovarian folliculogenesis in the mouse. Dev Biol 2001; 233(2):258-70.
Adashi E, Leung PCK. The Ovary: Comprehensive Endocrinology. Raven New York. 1993.
Ahmed N, Riley C, Rice G, Quinn M. Role of integrin receptors for fibronectin, collagen and laminin in the regulation of ovarian carcinoma functions in response to a matrix microenviroment. Clin Exp Metastasis 2005; 22(5):391-402.
Assou S, Anahory T, Pantesco V, Le Corrour T, Pellestor F, Lein B, Reyftmann L, Dechaud H, De Vos J, Hamamah S. Cumulus-oocyte complex gene-expression profile. Human Reprod 2006; 21(7):1705-1719.
Berkholtz CB, Lai BE, Woodruff TK, Shea LD. Distribution of extracellular matrix proteins type I collagen, type IV collagen, fibronectin and laminin in mouse folliculogenesis.
Histchem Cell Biol 2006a; 126(59):583-92.
Berkholtz CB, Shea LD, Woodruff TK. Extracellular matrix functions in follicle maturation. Semin Repro Med 2006b; 24(4):262-9.
Burns KH, Owens GE, Fernández JM, Nilson JH, Matzuk MM. Characterization of integrin expression in the mouse ovary. Biol Reprod 2002; 67(3):743-51.
Campbell S, Swann HR, Seif MW, Kimber SJ, Aplin JD. Cell adhesion molecules on the oocyte and preimplantation human embryo. Human Reprod 1995; 10(6):1571-8.
Clavero A, Castilla JA, Martínez l, Mendoza N, Fontes J, Maldonado V. Expression of integrin fraction and adhesion molecules on human granulosa cells and its relation with oocyte maturity and follicular steroidogenesis. J Assist Repro Genet 2004; 21(6):187-95.
Curry TE, Smith MF. Impact of extracellular matrix remodelling on ovulation and the follicle-luteal transition. Semin Reprod Med 2006; 24(4):228-41.
Driancourt MA, Reynaud K, Cortvrindt R, Smitz J. Roles of KIT and KIT LIGAND in ovarian function. Reproduction 2000; 5:143-52.
Evans WH, Martin PEM. Lighting up gap junction channels in a flash. BioEssays 2002; 24:876-80.
Fortune JE, Cishman RA, Kito WS. The primordial to primary follicle transition. Mol Cell Endocrinol 2000 ; 163:53-60,
Fujiwara H, Maeda M, Honda T, Yamada S, Ueda M, Kanzaki H, Suginami H, Mori T. Granulosa cells express integrin alpha 6: posible involvement of integrin alpha 6 in folliculogenesis. Horm Res 1996; 46(1):24-30.
Gittens JE, Barr KJ, Vanderhyden BC, Kidder GM. Interplay between paracrine signaling and gap junctional communication in ovarian follicles. J Cell Sci 2005a; 118(1):5071-8.
Gittens JE, Kidder GM. Differential contributions of connexin 37 and connexin 43 to oogenesis revealed in chimeric reaggregated mouse ovaries. J Cell Sci 2005b; 118(21):5071-8.
Goodenough DA, Simon AM, Paul DL. Gap Junctional intercellular communication in the mouse ovarian follicle. Novartis Found Symp 1999; 219:226-35.
Granot I, Dekel N. The ovarian gap junction protein connexion 43: regulation by gonadotropins. Trends Endocrinol Metab 2002; 13(7):310-13.
Guigon CJ, Magre SContribution of Germ cells to the differentiation and maturation of the ovary: insights from models of germ cell depletion. Biol Reprod . 2006; 74:450-58.
Hernández ER, Hurwitz A, Botero L, Ricciarelli E, Werner H, Roberts CT Jr, LeRoith D, Adashi EY. Insulinlike growth factor receptor gene expression in the rat ovary: divergent
regulation of distinct receptor species. Mol Endocrinol 1991; 5(12):1799-805.
Hernández ER, Hurwitz A, Vera A, Pellicer A, Adashi EY, LeRoith D, Roberts CTJr. Expression of the genes encoding the insulin-like growth factors and their receptors in the human ovary. J Clin Endocrinol Metab 1992; 74(2):419-25.
Honda T, Fujiwara H, Ueda M, Maeda M, Mori T. Integrin alpha 6 is a differentiation antigen of human granulosa cells. J Clin Endocrinol Metab 1995; 80(10):2899-905.
Hutt KJ, Mc Laughlin EA, Holland MK. Kit/Kit ligand in mammalian oogenesis and folliculogenesis: roles in rabbit and murine ovarian follicule activation and oocyte growth. Biol Reprod 2006; 75(3):421-3.
Irving-Rodgers HF, Rodgers RJ. Extracellular matrix in ovarian follicular development and disease. Cell Tissue Res 2005; 322(1):89-98.
Irving-Rodgers HF, Rodgers RJ. Extracellular matrix of the developing ovarian follicle. Semin Reprod Med 2006; 24(4):195-203.
Juneja SC, Barr KJ, Enders GC, Kidder GM. Defects in the germ line and gonads of mice lacking connexin 43. Biol Reprod 1999; 60(5):1263-70.
Kidder GM, Mhawi AA. Gap junctions and ovarian folliculogenesis. Reproduction 2002; 123(5):613-20.
Kocabas AM, Crosby J, Ross PJ, Out HH, Beyhan Z, Can H, Tam WL, Rosa GJ, Halgren RG, Lim B, Fernández E, Cibelli JB. The transcriptome of human oocytes. Proc Natl Acad Sci USA 2006; 103(38):14027-32.
Larue L, Ohsugi M, Hirchenhain J, Kemler R. E-cadherin null mutant embryos fail to form a trophectoderm epithelium. Proc Natl Acad Sci USA 1996; 91(17):8263-7.
Lei L, Zhang H, Jin S, Wang F, Fu M, Wang H, Xia G. Stage specific germ-somatic cell interaction directs the primordial folliculogenesis in mouse fetal ovaries. J Cell Physiol 2006; 208(3):640-7.
Lu DP, Tian L, O´Neill C, King NJ. Regulation of cellular adhesion molecule expression in murine oocytes, peri-implantation and post-implantation embryos. Cell Res 2002; 12(5-6):373-83.
Machell NH, Blaschuk OW, Farookhi R. Developmental expression and distribution of N- and E-cadherin in rat ovary. Biol Reprod. 2000; 63(3):797-804.
Machell NH, Farookhi R. E- and Ncadherin expression and distribution during luteinization in the rat ovary. Reproduction 2003; 125(6):791- 800.
Matzuk MM, Burns KH, Viveiros MM, Eppig JJ. Intercellular Communication in the Mammalian Ovary: Oocytes Carry the conversation. Science 2002; 296:2178-180.
Miyoshi T, Otsuka F, Suzufki J; Takeda M, Inagaki K, Kano Y, Otani H, Mimura Y, Ogura T, Makino H. Mutual Regulation of Follicle- Stimulating Hormone Signaling and Bone Morphogenetic Protein System in Human Granulosa Cells. Biol Reprod 2006; 74(6):1073-82.
Nakamura K, Fujiwara H, Higuchi T, Honda T, Nakayama T, Kataoka N, Fujita K, Ueda M, Maeda M, Mori T. Integrin alpha6 is involved in follicular growth in mice. Biochem Biophys Res Commun 1997; 235(3):524-8.
Pangas SA, Woodruff TK. Activin signal transduction pathways. Trends Endocrinol Metab 2000; 11(8):309-14.
Pate BJ, White KL, Winger QA, Rickords LF, Aston KI, Sessons BR, Li GP, Campbell KD, Weimer B, Bunch TD. Specific integrin subunits in bovine oocytes, including novel sequences for alpha 6 and beta 3 subunits. Mol Reprod Dev 2006; 12.
Peramo B, Ricciarelli E, Hernández ER. Regulación intraovárica de la foliculogénesis. Factores de crecimiento. Reproducción Humana. McGraw-Hill. Interamericana; 2002.
Russel DL, Salustri A. Extracellular matrix of the cumulus-oocyte complex. Semin Reprod Med 2006; 24(4):217-27.
Sato E, Yokoo M. Morphological and biochemical dynamics of porcine cumulus-oocyte complexes: role of cumulus expansion in oocyte maturation. Ital J Anat Embryol 2005;
110(2):205-17.
Schuldt A. Cadherins reach out. Nat Cell Biol 2005; 7(12):1066.
Shimada M, Terada T. FSH and LH induce progesterone production and progesterone receptor synthesis in cumulus cells: a requirement for meiotic resumption in porcine oocytes. Mol Hum Reprod 2002; 8(7):612-8.
Shozu M, Minami N, Yokoyama H, Inoue M, Karihara H, Matsushima K, Kuno K. ADAMTS-1 is involved in normal follicular development, ovulatory process and organization of the medullary vascular network in the ovary. J Mol Endocrinol 2005; 35(2):343-55.
Simon AM, Mc Whorter AR. Vascular abnormalities in mice lacking the endothelial gap junction proteins connexin 37 and connexin 40. Dev Biol 2002; 251(2):206-20.
Simon AM, Chen H, Jackson CL. Cx37 and Cx43 localize in zona pellucida in mouse ovarian follicles. Cell Commun Adhes 2006; 13(1- 2):61-77.
Takao Y, Fujiwara H, Yamada S, Hirano T, Maeda M, Fujii S, Ueda M. CD9 is expressed on the cell surface of human granulosa cells and associated with alpha6beta1. Mol Hum Reprod 1999; 5(4):303-10.
Teilmann SC. Differential expression and localization of connexin-37 and connexin-43 in follicles of different stages in the 4-week-old mouse ovary. Mol Cell Endocrinol 2005; 234(1-2):27-35.
Thomas FH, Vanderhyden BC. Oocyte-granulosa cell interactions during mouse follicular development: regulation of kit ligand expression and its role in oocyte growth. Reprod Biol Endocrinol 2006; 12(4):19.
Wright CS, Becker DL, Lin JS, Warner AE, Hardy K. Stage-specific and differential expression in the mouse ovary: connexin-specific roles in follicular regulation. Reproduction 2001; 121(1):77-88.
Yogo K, Ogawa T, Akiyama M, Ishida N, Takeya T. Identification and functional analysis of novel phosphorylation sites in Cx43 in rat primary granulosa cells. FEBS Lett 2002; 531(2):132-6.
Yogo K, Ogawa T, Akiyama M, Ishida- Kitagawa N, Sasada H, Sato E, Takeya T. PKA implicated in the phosphorylation of Cx43 induced by stimulation with FSH in rat granulose cells. J Reprod Dev 2006;52(3):321-8
Yoon S, Kim K, Chung H, Choi D, Lee W, Cha K, Lee K. Gene expression profiling of early follicular development in primordial, primay, and secondary follicles. Fert Ster 2006; 85 (1): 193-203.
Ziv S, Rufas O, Shalgi .R .Cadherins expression during gamete maturation and fertilization in the rat. Mol Reprod Dev 2002 ; 62 (4): 547-56.