MULTINUCLEACIÓN Y DESARROLLO EMBRIONARIO
Lucía Hortal, María José de los Santos
IVI Valencia
email: luciahortal@gmail.com
Fecha recepción: 3 Abril 2012
Fecha aceptación: 16 Septiembre 2012
La Multinucleación (MN) definida como la presencia de dos o más núcleos en una o más blastómeras del preembrión humano, aparece entre un 17% y un 69% de los casos dependiendo de series y autores. Su presencia parece estar asociada a diferentes condicionantes exógenos como pueden ser protocolos de estimulación, así como a otras variables morfológicas embrionarias como tamaño de las blastómeras, fragmentación y cinética evolutiva. El objetivo de este estudio es hacer una revisión de los factores que influyen en su aparición y la correlación que presenta con otras características morfológicas, así como examinar su repercusión en el desarrollo embrionario, en las tasas de implantación y en la aparición de aneuploidías. Rev Asoc Est Biol Rep 2012; 17(2): 23-28.
INTRODUCCIÓN
La multinucleación (MN), definida como la presencia de dos o más núcleos en una o más blastómeras del embrión, se considera como una característica patológica y de mal pronóstico reproductivo. Aparece indiferentemente en condiciones in vitro e in vivo, caracterizándose por no presentar una distribución normal en la población sino apareciendo como un suceso dependiente y vinculado a cada paciente (Van Royen et al., 2003).
La frecuencia de MN varía en un rango del 17 al 69% en embriones humanos, viéndose incrementada cuando coexiste con otras anormalidades morfológicas (Kligman et al., 1996; Balakier et al., 1997; Jackson et al., 1998; Pelinck et al., 2010; Van Royen et al., 2003). La frecuencia con la que se encuentran blastómeras con MN en embriones en D+2/D+3 procedentes de técnicas de fecundación in vitro, oscila del 14,3% al 74% (Gil et al., 2007)aunquelamayoríadelas blastómeras multinucleadas parecen registrarse en D+2. La incidencia decrece del día 2 al día 3 (26,2% vs 11,4%) (Yakin et al., 2005).
La valoración de parámetros morfológicos (número de células, simetría, fragmentación, MN,…), parece ser la práctica más utilizada para seleccionar el mejor embrión a transferir. La MN es aceptada como un criterio morfológico relacionado con baja calidad embrionaria y se utiliza para decidir qué embrión resulta más adecuado a la hora de realizar una transferencia o una criopreservación. La lenta capacidad de desarrollo o el bloqueo mostrado por algunos embriones está relacionado con la presenciadeMN(Hardyetal.,1993), pero además diversos estudios han investigado su influencia negativa en las tasas de implantación y en la aparición de aneuploidías, por ello la utilización de la MN como indicador de calidad en los laboratorios de FIV parece más que justificada (Hesters et al., 2008; Van Royen et al., 2001; Van Royen et al., 2003).
La presencia de MN no siempre es fácil de observar; numerosos factores como la granulosidad del citoplasma, la superposición de blastómeras, etcétera, pueden influir en la valoración de ésta. Debido a la importancia que tiene su detección, diversos autores han aportado el análisis de otros parámetros que pueden ayudar a localizarla. Hardarson et al., 2001, indicaba en su estudio un incremento en las tasas de MN y de implantación en embriones con tamaño desigual de blastómeras y señalaba cómo dichas blastómeras se veían afectadas por un mayor número de anomalías cromosómicas numéricas.
Estudios posteriores encontraban que los blastómeros con tamaño más grande presentaban mayores tasas de MN; el volumen medio registrado en las blastómeras no multinucleadas fue significativamente más pequeño que en las blastómeras multinucleadas (Hnida et al., 2004). Con la intención de aumentar la precisión en la medición del tamaño de las blastómeras se ha utilizado el análisis computerizado para valorar la calidad embrionaria, en cuanto a la evaluación de la fragmentación y la MN.
MATERIAL Y MÉTODOS
El diseño que se realizó se encuadra dentro de una Revisión Sistemática, en la que se pretendió recopilar, evaluar y obtener conclusiones de las investigaciones realizadas acerca de la influencia de la MN en el desarrollo embrionario. La búsqueda bibliográfica se realizó en dos fases. La primera automatizada a través de la base de datos MEDLINE. Se establecieron criterios de búsqueda basados en palabras clave (MN, aneuploidía, desarrollo embrionario, tasa implantación, selección embrionaria, patrón de división, blastocisto) y limitado en un principio a los 10 últimos años. Se seleccionó como idioma preferente el inglés y el español. En la segunda fase, se obtuvieron estudios más relevantes de forma manual a partir de la lectura de los estudios seleccionados en la primera fase. Se realizó una búsqueda de los últimos boletines presentados en congresos publicados en la revista ASEBIR, con el fin de obtener la última información sobre este tema en el ámbito nacional.
RESULTADOS
Dada la influencia negativa que la MN tiene sobre el desarrollo embrionario, los objetivos de la presente revisión bibliográfica son recopilar, sintetizar y evaluar los posibles orígenes y factores que intervienen en la aparición de la MN, analizando la influencia negativa de ésta en el patrón de división embrionario y en las tasas de implantación, y su correlación con las anormalidades cromosómicas.
FACTORES Y CARACTERÍSTICAS MORFOLÓGICAS RELACIONADAS CON LA APARICIÓN DE MN
La relevancia que tiene detectar la MN en las técnicas de selección, ha llevado a estudiar los posibles factores que pueden influenciar su aparición, con el fin de poder disminuir su presencia en los laboratorios de FIV. Se han abordado diferentes factores en el proceso de FIV que podrían influir en la aparición de la MN. A continuación, se describirán los factores más influyentes en su aparición, como el tipo de estimulación (principalmente las dosis, el tipo de gonadotropina utilizada y la cantidad de ovocitos recuperados), la maduración ovocitaria posestimulación y las condiciones del cultivo embrionario. Por otro lado, se analizará la aparición de la MN en relación con las otras variables morfológicas, como fragmentación, patrón pronuclear, tamaño de las blastómeras.
POSIBLES FACTORES QUE TIENEN INFLUENCIA EN LA APARICIÓN DE LA MN
Influencia de la estimulación
Varios trabajos hasta la fecha han mostrado que el tipo de estimulación influye en la aparición de blastómeras multinucleadas en los embriones. Su existencia parece estar asociada con ciclos de estimulación ovárica en los cuales los ovarios responden de manera masiva por una alta sensibilidad a la terapia con gonadotropinas (Jackson et al., 1998; Van Royen et al., 2003; Brossa et al., 2008).
En un estudio retrospectivo, Van Royen et al. (2003), analizaron diferentes parámetros dentro de la estimulación que podrían estar influyendo en la aparición de la MN. El estudio retrospectivo incluyó 10.388 embriones derivados de 1395 ciclos consecutivos de FIV / ICSI en 700 pacientes. Las pacientes pertenecientes a un rango de edad comprendido entre 20 – 45 años, fueron estimuladas con un protocolo largo. Se utilizaron tres gonadotropinas diferentes, las cuales fueron administradas en distintas dosis según requerimientos de la paciente. Los ovocitos obtenidos fueron fecundados mediante FIV o ICSI. Los embriones fueron evaluados en día 2 y día 3 en función de tres parámetros; fragmentación, número de blastómeras y número de blastómeras multinucleadas. Tras el análisis de los resultados se observó que el número de ovocitos recuperados y la dosis de gonadotropina utilizada, tuvieron un impacto en la MN. Concretamente, los ciclos en los que se obtuvieron 9 ovocitos o menos, presentaron embriones con menor MN que cuando se obtuvieron 10 o más ovocitos por punción (29,7% vs 34,5%; RR = 1.16, 95% CI = 1.08±1.25). Las estimulaciones que necesitaron una gran dosis de gonadotropina también fueron asociadas con un incremento en la MN (≥ 2400 UI, 34,6%; < 2400 UI, 32,0%; RR =1.08, 95% CI = 1.02±1.14). No se observaron diferencias significativas en cuanto a la duración o el tipo de gonadotropina usada, edad materna o técnica utilizada. Estos datos fueron analizados en otro estudio retrospectivo posterior, teniendo en cuenta únicamente la dosis de FSH utilizada y no pareció observarse mayor MN. Sin embargo, sí observaron un aumento de la incidencia en aquellos ciclos con dosis > 2400 UI de FSH y en los que se recuperaban más de 10 ovocitos (Brossa et al., 2008).
En los estudios anteriormente citados (Brossa et al., 2008; Van Royen et al., 2003) no se encontraron diferencias significativas en referencia al tipo de gonadotropina utilizada. Sin embargo, Vlaisavljevic et al. (2006) presentaban unos resultados que apuntaban a un menor porcentaje de embriones con MN en las pacientes tratadas con distinto tipo de gonadotropina, hCG versus FSH (30,0% vs 66,7; P= 0,0123).
Otras de las variables asociadas a la estimulación, como por ejemplo el tipo de supresión hipofisiaria, también se ha visto que podría tener repercusión en la MN; por ejemplo en un estudio realizado por De Cassia Savio et al. (2010), se encontraron diferencias que podrían influir en el aumento de la MN. Los resultados mostraron cómo la supresión con agonistas de la GnRH provocaba la obtención de embriones con mayores tasas de MN, en comparación con el uso de antagonistas (33,6% vs 28,3%, P= 0,046). Estos autores consideraron que la selección de folículos con agonistas era menos exigente, resultando en una mayor recuperación de ovocitos y en la obtención de menores tasas de embarazo a partir de embriones con MN (16,6%, P= 0,07).
Maduración incompleta de ovocitos
Se ha demostrado que los ovocitos madurados in vitro son sensibles a la edad posmaduración y que esto puede resultar en un alto grado de anormalidades pronucleares, así como en un incremento en las tasas de MN (Balakier et al., 2004).
Los estudios basados en el análisis de ovocitos madurados in vitro han aportado datos relevantes sobre la influencia que tiene en la aparición de embriones multinucleados. En uno de ellos, los ovocitos recuperados en MII formaron el grupo control, el otro grupo lo constituyeron ovocitos en MI sometidos a cultivo hasta alcanzar MII. Todos los ovocitos fueron fecundados mediante ICSI y evaluados posteriormente. Entre los resultados obtenidos, se pudieron observar diferencias significativas con respecto a la presencia de MN en ambos grupos, siendo casi el doble en los embriones derivados de ovocitos madurados in vitro de MI a MII que en el grupo control (23 – 13%; P< 0,0001) (Tabla I).
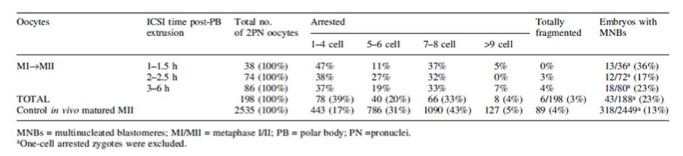
Por otro lado, tal y como muestra Nogueira et al. (2000) en su estudio sobre el estatus nuclear y citogenético de embriones derivados de ovocitos madurados in vitro, la MN fue un factor encontrado en un alto porcentaje de estos embriones en día 2 y en día 3 (16% binucleados; 26% multinucleados; 33% anucleados). Además, tan sólo el 60% de estos embriones mostraron una buena morfología y el 44% quedaron bloqueados en su primera división mitótica. Los autores apuntaron la posibilidad de la falta de maduración del citoplasma y de cómo esta deficiencia podría no ser rescatada por el espermatozoide, lo que supondría defectos en el huso mitótico durante la división. Esto afectaría a un óptimo desarrollo embrionario y podría explicar la baja tasa de implantación que presentan estos embriones.
Condiciones de cultivo
Otro factor asociado con la formación de la MN son las posibles variaciones de temperatura o condiciones subóptimas que se puedan dar durante el cultivo del embrión, así como la falta de componentes moleculares que desencadenan la división citoplasmática. Esto podría tener un grave efecto en la función del citoesqueleto, provocando divisiones celulares anómalas (Pelinck et al., 1998; Hardy et al., 1993). Sin embargo hay muy pocos estudios publicados que aborden el tema específico de la MN con este tema en concreto.
RELACIÓN DE LA MN CON OTRAS CARACTERÍSTICAS MORFOLÓGICAS
Coexistencia con fragmentación
Al parecer, la presencia de MN, aunque se puede presentar de forma aislada, tiene cierta asociación con otras variables de carácter morfológico; en este sentido se ha registrado una mayor incidencia de MN en embriones con una alta fragmentación (Hardarson et al., 2001; Vlaisavljevic et al., 2006). En su análisis Van Royen et al. (2003), encontraron diferencias significativas entre los diferentes grados de fragmentación y el empeoramiento de la MN. Los grados de fragmentación fueron definidos en función del porcentaje de fragmentos anucleados existentes en cada embrión (F1:≤10%; F2: 10-20%; F3: 20-30%). Embriones con un menor grado de fragmentación (F1) mostraban menor MN que embriones con fragmentaciones tipo 2 (F2) o tipo 3 (F3). Esto podría explicarse por una dificultad de observación debida a la existencia de gran cantidad de fragmentos.
En otro estudio del mismo autor, se pudo comprobar que la tasa de implantación para embriones con un porcentaje de fragmentación mayor o igual al 20%, y queademáspresentabanMN,fuemenor o igual al 3% (Van Royen et al., 2001).
Correlación con el patrón pronuclear
Otro parámetro relacionado con la MN es el llamado score pronuclear. Consiste en la evaluación de la morfología pronuclear del zigoto y en la determinación del número y distribución de los cuerpos precursores nucleolares (NPB). Es utilizado actualmente en la valoración de la calidad embrionaria. Su relación con la MNfueevaluadaenembrionesdedía2y día 3, en un estudio realizado por Gámiz et al. (2003). Las tasas de MN obtenidas fueron similares en embriones de día 2.
Sin embargo, su porcentaje se duplicó cuando se observó en embriones de día 3 procedentes de zigotos con diferente tamaño pronuclear frente a embriones con el mismo tamaño de pronúcleos, pasando del 5,4% al 10,9%. Tamaños desiguales en los pronúcleos, así como grandes distancias entre ellos, están asociados con elevadas incidencias en la MN y con bajas tasas de desarrollo.
CONSECUENCIAS DE LA MN EN LA VIABILIDAD EMBRIONARIA
Influencia en el patrón de división
Diversos estudios han analizado la relación existente entre la presencia de MN y el patrón de división. Según Van Royen et al., un óptimo patrón de división tiende a presentarse en embriones mononucleados, con una mínima incidencia de MN (Van Royen et al., 2001; Van Royen et al., 2003). El impacto en la tasa de división cuando los embriones presentanMN,sepresentóenotro análisis retrospectivo en el cual se mostraba cómo disminuía del 91,6% en embriones sin MN al 56% – 36,4%, si el embrión contenía una o más blastómeras MN, respectivamente (P< 0,001) (Yakin et al., 2005).
Hardyetal.(1993)apuntaban que la disminución en la tasa de división podría deberse a la existencia de blastómeras con cromosomas anormales. La presencia de núcleos anormales favorece un arresto en la división embrionaria provocando que estas células no puedan realizar un desarrollo competente (Hardy et al., 1993; Yakin et al., 2005).
La implicación de la MN, en referencia a la influencia sobre el patrón de división y el desarrollo del embrión, también se encuentra al evaluar la tasa de formación de blastocisto. Yakin et al. (2005) registraban una disminución en las tasas de formación de blastocisto enembrionesmultinucleados(51%vs 11,4%-6,55; P< 0,0001). La calidad del blastocisto también se veía comprometida.
La evaluación de la MN y el desarrollo a blastocisto fue estudiada también por Alikani et al. (2000) anotando que tan sólo el 15,9% de los embriones que contenían MN en día 2 y en día 3 llegaron a formar blastocistos normales, frente a una tasa del 31,9% en embriones no multinucleados. La competencia de estos embriones para compactar fue de un 30,6% frente a un 47% en los embriones sin MN. De la misma manera, los valores obtenidos frente a la capacidad de cavitación fueron de un 27,5% frente a un 45,9%, respectivamente.
MN- Anormalidades cromosómicas
La gran mayoría de estudios realizados sobre el estatus cromosómico de las blastómeras multinucleadas muestran que son cromosómicamente anormales (Ambroggio et al., 2011; Kligman et al., 1996; Nogueira et al., 2000; Hardarson et al., 2001). Dichas aportaciones afianzan la idea de que estos embriones son, en su mayoría, incompetentes para producir fetos viables, obteniéndose mayoresfallosdedesarrolloenellos; por tanto, la tendencia general es a evitar su transferencia, a menos que no haya otro embrión con mejor pronóstico.
La aparición de estas anormalidades estáasociadaconasincroníasentrelos procesosdecariocinesisycitocinesis durante el desarrollo del embrión. Se han propuesto hipótesis como la incompleta división citoplasmática pero completa división del núcleo (Hardy et al., 1993); posibles errores en la segregación de cromosomas, fragmentación parcial del núcleo, organizaciones anómalas del huso mitótico que pueden acabar con errores de empaquetamiento durante la migración de los cromosomas en la anafase (Kligman et al., 1996).
En un análisis con hibridación in situ por fluorescencia (FISH) realizado por Kligman et al. (1996) se mostraron unos resultados que apuntaban a un aumento de anormalidades cromosómicas en embriones multinucleados frente a embriones mononucleados (76,6% vs 50,9%; P <0,001). Además, manifestaban que la cantidad de células MN en un embrión, la aparición de la MN en D2 o D3, los embriones con un desarrollo lento o normal, y embriones obtenidos a partir de ICSI o inseminados, no presentaban diferencias en cuanto a la incidencia de anormalidades cromosómicas.
Algunos autores han sugerido que la MN podría ser un fenómeno temporal y reversible sin implicar siempre anormalidades cromosómicas. (Gil et al., 2007; Staessen et al., 1998). De hecho, Kligman et al. (1996) propusieron a partir de los resultados que obtuvieron la existencia de dos patrones diferentes de MN. Uno de ellos es el que se produciría en D2 y D3 del desarrollo, el cual produciría mayoritariamente embriones con anormalidades cromosómicas, y el otro, se encontraría en D4 o posterior de desarrollo, y resultaría en embriones cromosómicamente normales.
La desaparición de la MN en etapas posteriores del desarrollo se ha intentado explicar mediante distintas hipótesis. Una de las hipótesis propuestas, es la posibilidad de inestabilidad en el número de núcleos y la posterior fusión de éstos, dando como resultado la desaparición de la MN en estadios posteriores. Otras vertientes, exponen la posibilidad de una reanudación posterior de la citocinesis, el desplazamiento al trofoectodermo de las células cromosómicamente anormales o incluso falloseneldiagnósticodelaMN(Gilet al., 2007).
La presencia de MN puede estar asociada a mosaicismo, como así lo demostró un estudio de Magli y colaboradores en el que se observó que un 92% de los embriones con MN resultaron ser mosaicos (Magli et al., 2001).
Un estudio publicado recientemente se centró en analizar, en embriones de día 2 mediante biopsia y PGD en día 3, el impacto de la MN en las tasas de aneuploidía y en las tasas de implantación. Los cromosomas analizados fueron X, Y, 13, 15, 16, 18, 21, 22 mediante hibridación in situ con fluorescencia (FISH). Los embriones con blastómeras multinucleadas tendían a presentar una mayor incidencia de aneuploidía en comparación con los embriones que mostraban únicamente blastómeras mononucleadas (85% vs 78%; RR 0,92, IC 95%: 0,84-1,0) (Ambroggio et al., 2011). Aunque los resultados no fueron estadísticamente significativos (RR 0,92, IC 95% 0.84- 1.00), los autores consideraron su estudio lo suficientemente consistente para detectar una tendencia al incremento en las tasas de aneuploidías del 7%, entre embriones con MN y sin MN. Ningún embrión con MN consiguió implantar.
A pesar de que una gran cantidad de embriones con MN contienen anormalidades cromosómicas, no está claro si dichas aberraciones están concentradas únicamente en las células multinucleadas o son compartidas por el resto de las células. Esto podría tener implicaciones en cuanto al desarrollo, ya que un embrión en el que sus blastómeras MN detuvieran su desarrollo, aún podría desarrollarse normalmente a partir del resto. Por el contrario, embriones con muchas blastómeras MN podrían ver perjudicado su desarrollo. En este proceso de división también influiría el momento de aparición de la MN, ya que si apareciera durante las primeras divisiones embrionarias, todas las células del embrión podrían verse afectadas (Kligman et al., 1996).
Viabilidad de embriones MN e influencia en la capacidad de implantación
Dada la asociación de la MN con anomalías cromosómicas, así como su asociación con el ritmo de división o la elevada tasa de fragmentación y la asimetría, es de esperar que los embriones multinucleados tengan un menor potencial biológico.
Las bajas tasas de implantación derivadas de la presencia de MN en los embriones, hacen de la MN un factor altamente discriminatorio. Diversos artículos resaltan la obtención de tasas de implantación más altas realizando transferencias de embriones sin MN (15% vs 5%; P=0,042) (Jackson et al., 1998), e incluso se apuntan tasas de implantación comprometidas cuando la MN se presenta asociada a la fragmentación (Van Royen et al., 2001). A pesar de la influencia que tiene la MN sobre la implantación, algunos autores han expuesto que el número de blastómeras que conforma un embrión interfiere más que la presencia de MN (Pelinck et al., 2010).
Algunos autores han encontrado una mejora en las tasas de implantación cuando las transferencias son hechas sólo con embriones mononucleados a cuando éstas son mixtas (mononucleadas y multinucleadas), (11,3% – 6%; P= 0.003) (Pelinck et al., 1998). Gil et al. (2007) no encontraron diferencias significativas en los resultados obtenidos en su análisis con respecto a este dato, pues aunque la tasa de implantación fue mayor en las transferencias mononucleadas (19%) que en las mixtas (15,8%), las diferenciasnofueronestadísticamente significativas. Esto podría deberse a esa corrección de la MN citada anteriormente.
La baja tasa de implantación de embriones con MN supone una disminución de recién nacidos vivos. Este hecho podría correlacionarse con la existencia de aneuploidías y con la influencia que la MN tiene en el patrón de división (Alikani et al., 2000; Balakier et al., 1997; Kligman et al., 1996; Pelinck et al., 1998; Staessen et al., 1998; Yakin et al., 2005).
Teniendo en cuenta las repercusiones que la MN tiene sobre el patrón de división, la aparición de aneuploidías y lastasasdeimplantación,parecelógico que estos embriones no se utilicen para realizar transferencias, a no ser que no haya otros embriones para elegir.
Diversos estudios han demostrado la existencia de recién nacidos sanos a partirdetransferenciasconembriones con MN. El estudio de Balakier et al. (1997) indicaba la transferencia de 19 embriones con MN, de los cuales tan solo 4 implantaron (21%), dos de ellos acabaron en aborto espontáneo y dos finalizaron con el nacimiento de dos recién nacidos sanos. Los embriones que llegaron a término estaban formados por blastómeras multinucleadas y mononucleadas, considerando que estos embriones pudieran sufrir un bloqueo de sus blastómeras con MN y un desarrollo a partir de las mononucleadas. Esto podría indicar la totipotencia blastomérica, es decir, la necesidad de tener sólo una célula mononucleada para realizar un desarrollo embrionario completo en estadio de 2 células.
En la misma línea, Van Royen et al. (2003) obtenían unos resultados en los que se apuntaba el nacimiento de un niño sano a partir de una transferencia simple de un embrión con MN. Pelinck et al. (1998) también mostraron resultados de recién nacidos vivos a partir de embriones con MN, en los cuales de 8 transferencias realizadas, tan sólo se obtuvo el nacimiento de un niño sano. También Jackson et al. (1998) mostraron 25 ciclos en los que todas las transferencias tenían embriones con al menos una blastómera con MN. Implantaron 3 embriones, cuyo final fue un aborto espontáneo y dos gemelos sanos.
CONCLUSIONES
A la vista de las repercusiones que la MN tiene sobre las tasas de implantación, la aparición de aneuploidías y la viabilidad embrionaria, parece clara la necesidad de realizar esfuerzos en los laboratorios de embriología clínica centrados en mejorar su detección y conocer su repercusión biológica en el embrión como fenómeno aislado. Ahora bien, dichos esfuerzos no sólo deberían ir dirigidos a realizar una buena evaluación morfológica del embrión, sino que habrían de centrarse en controlar todos aquellos factores externos que tienen influencia en la aparición de dicha patología.
Referencias
Alikani M, Calderon G, Tomkin G, Garrisi J, Kokot M, Cohen J. Cleavage anomalies in early human embryos and survival after prolonged culture in-vitro. Hum Reprod 2000;15: 2634-2643.
Ambroggio J, Gindoff PR, Dayal MB, Khaldi R, Peak D, Frankfurter D, et al. Multinucleation of a sibling blastomere on day 2 suggests unsuitability for embryo transfer in IVF- preimplantation genetic screening cycles. Fertil Steril 2011; 96: 856-859.
Balakier H, Cadesky K. The frequency and developmental capability of human embryos containing multinucleated blastomeres. Hum Reprod 1997; 12: 800-804.
Balakier H, Sojecki A, Motamedi G, Librach C. Time-dependent capability of human oocytes for activation and pronuclear formation during metaphase II arrest. Hum Reprod 2004; 19: 982-987.
Brossa M, Blanch X, Antich M. Incidencia de multinucleación en IVF-ICSI. ASEBIR 2008; 16-18.
De Cassia Savio FR., Souza SA., Paes De Almeida Ferreira Braga, Iaconelli A Jr, Borges E Jr. Blastomere multinucleation: Contributing factors and effects on embryo development and clinical outcome. Hum Fertil 2010; 13: 143-150.
Gamiz P, Rubio C, de los Santos MJ, Mercader A, Simon C, Remohi J, et al. The effect of pronuclear morphology on early development and chromosomal abnormalities in cleavage-stage embryos. Hum Reprod 2003; 18: 2413-2419.
Gil M, D’Ommar G, Poo ME, Sosa A, Piras M, Piras R, et al. Insights on blastomere nuclearity. J Assist Reprod Genet 2007; 24: 17-22.
Hardarson T, Hanson C, Sjogren A, Lundin K. Human embryos with unevenly sized blastomeres have lower pregnancy and implantation rates: indications for aneuploidy and multinucleation. Hum Reprod 2001; 16: 313-318.
Hardy K, Winston RM, Handyside AH. Binucleate blastomeres in preimplantation human embryos in vitro: failure of cytokinesis during early cleavage. J Reprod Fertil 1993; 98: 549-558.
Hesters L, Prisant N, Fanchin R, Mendez Lozano DH, Feyereisen E, Frydman R, et al. Impact of early cleaved zygote morphology on embryo development and in vitro fertilization-embryo transfer outcome: a prospective study. Fertil Steril 2008; 89: 1677-1684.
Hnida C, Engenheiro E, Ziebe S. Computer- controlled, multilevel, morphometric analysis of blastomere size as biomarker of fragmentation and multinuclearity in human embryos. Hum Reprod 2004; 19: 288-293.
Jackson KV, Ginsburg ES, Hornstein MD, Rein MS, Clarke RN. Multinucleation in normally fertilized embryos is associated with an accelerated ovulation induction response and lower implantation and pregnancy rates in in vitro fertilization-embryo transfer cycles. Fertil Steril 1998; 70: 60-66.
Kligman I, Benadiva C, Alikani M, Munne S. The presence of multinucleated blastomeres in human embryos is correlated with chromosomal abnormalities. Hum Reprod 1996; 11: 1492-1498.
Magli MC; Gianaroli L, Ferraretti AP. Chromosomal abnormalities in embryo MolCell Endocrinol 2001; 183: S29-34.
Nogueira D, Staessen C, Van de Velde H, Van Steirteghem A. Nuclear status and cytogenetics of embryos derived from in vitro-matured oocytes. Fertil Steril 2000; 74: 295-298.
Pelinck MJ, De Vos M, Dekens M, Van der Elst J, De Sutter P, Dhont M. Embryos cultured in vitro with multinucleated blastomeres have poor implantation potential in human in- vitro fertilization and intracytoplasmic sperm injection. Hum.Reprod 1998; 13: 960-963.
Pelinck MJ, Hoek A, Simons AH, Heineman MJ, van Echten-Arends J, Arts EG. Embryo quality and impact of specific embryo characteristics on ongoing implantation in unselected embryos derived from modified natural cycle in vitro fertilization. Fertil Steril 2010; 94: 527-534.
Staessen C, Van Steirteghem A. The genetic constitution of multinuclear blastomeres and their derivative daughter blastomeres. Hum.Reprod 1998; 13: 1625-1631.
Van Royen E, Mangelschots K, De Neubourg D, Laureys I, Ryckaert G, Gerris J. Calculating the implantation potential of day 3 embryos in women younger than 38 years of age: a new model. Hum.Reprod 2001; 16: 326-332.
Van Royen E, Mangelschots K, Vercruyssen M, De Neubourg D, Valkenburg M, Ryckaert G, et al. Multinucleation in cleavage stage embryos. Hum.Reprod 2003; 18: 1062-1069.
Vlaisavljevic V, Cizek-Sajko M, Kovac V. Multinucleation and cleavage of embryos derived from in vitro-matured oocytes. Fertil Steril 2006; 86: 487-489.
Yakin K, Balaban B, Urman B. Impact of the presence of one or more multinucleated blastomeres on the developmental potential of the embryo to the blastocyst stage. Fertil. Steril 2005; 83: 243-245.